1.本发明涉及竹林经营领域,具体涉及一种测算单株毛竹扩散能力及关键调节因子的方法。
背景技术:
2.毛竹(phyllostachys edulis)原产于我国亚热带地区,属于禾本科(gramineae)竹亚科(bambusoideae)刚竹属(phyllostachys),是典型的无性系植物,为单轴型散生竹类。竹养鞭、鞭生芽、芽育笋,笋成竹
……
周而复始,生生不息,逐年向四周扩散,对邻近森林群落结构组成、生物多样性、土壤理化性质等方面产生严重影响,受到中外学者高度关注并开展毛竹扩张方面的研究。然而缺乏对单株毛竹向周边扩散能力测算及关键抑制因子研究,而单株毛竹扩散是毛竹林向外扩张的源头,单株毛竹扩散能力及关键抑制因子的测算是摸清毛竹向外扩张的基础,为提出科学且有针对性的抑制技术提供依据。
3.关于毛竹扩散方面的研究,主要集中在毛竹扩张产生的影响,如对林地土壤肥力(吴家森,姜培坤,王祖良.天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响[j].江西农业大学学报,2008,30(4):689
‑
692)、凋落物水文功能(宋庆妮,杨清培,欧阳明,等.毛竹扩张的生态后效:凋落物水文功能评价[j].生态学杂志,2015,34(8):2281
‑
2287)、周边森林群落菌根系统(潘璐,牟溥,白尚斌,等.毛竹林扩张对周边森林群落菌根系统的影响[j].植物生态学报,2015,39(4):371
‑
382)等的影响;同时,根据毛竹鞭根状态分析,提出关于抑制毛竹林扩散的研究,如蔡亮等(蔡亮,张瑞霖,李春福,等.基于竹鞭状态分析的抑制毛竹林扩散的方法.东北林业大学学报,2003,31(5):68-70.)提出了挖沟、灌水的设想;另外,根据毛竹生物学特性和培育目的,国家林业局也发布了相关的技术规程《毛竹林丰产技术》(gb/t 20391
‑
2006)。综合上述毛竹相关研究,多是揭示毛竹扩张对其周边森林群落产生的影响,而单株毛竹扩散能力及关键调节因子的测算方法研究,未见报道。
技术实现要素:
[0004]
本发明提供了一种测算单株毛竹扩散能力及关键调节因子的方法,该方法较现有的人工开挖地下竹鞭计算单株毛竹扩散能力的方法更省力省时,效率明显提高,而且不会产生因开挖竹鞭产生对土壤结构、微生物的变化及影响。
[0005]
一种测算单株毛竹扩散能力及关键调节因子的方法,包括步骤:
[0006]
s1.选取有毛竹分布的林分样地;
[0007]
s2.在样地内标记各单株毛竹,标记满两年以标记的单株毛竹为圆心、10米为半径设置样圆,作为圆心的单株毛竹记作中心竹株,测定样圆内新增毛竹株数及各新增毛竹分别距中心竹株的距离,计算单株毛竹扩散能力指标;
[0008]
所述单株毛竹扩散能力指标的计算公式如下:
[0009]
[0010][0011][0012]
式中,r
i
为第i株中心竹株的扩散速率,d
ij
为第i株中心竹株与第j株新增毛竹的距离,a为样地调查间隔期2年,k
i
为第i株中心竹株10米半径样圆内的新增毛竹株数,j为第i株中心竹株10米半径样圆内新增的第j株毛竹,r1为样地内n株中心竹株的平均扩散速率即毛竹平均扩散速率,r2为单株中心竹株平均每年新增的毛竹株数;
[0013]
s3.确定样地林分内毛竹和/或木本植物的物种名称,测量得到样地林分内毛竹和/或木本植物的胸径和树高,并采用相邻网格法记录毛竹和/或木本植物的位置坐标(x、y);
[0014]
s4.根据所述物种名称、胸径、树高和位置坐标(x、y)测量数据,计算样地林分结构指标:林分密度、平均胸径、平均树高、物种丰富度指数、玛格列夫丰富度指数、香农
‑
维纳多样性指数、皮卢均匀度指数、树种简单混交度、多样性混交度、竞争指数和聚集指数;
[0015]
s5.将r1、r2对应记作y1、y2归为一组变量组y,将林分密度、平均胸径、平均树高、物种丰富度指数、玛格列夫丰富度指数、香农
‑
维纳多样性指数、皮卢均匀度指数、树种简单混交度、多样性混交度、竞争指数和聚集指数对应记作x1、x2、x3、x4、x5、x6、x7、x8、x9、x
10
、x
11
归为一组变量组x,经标准化后进行典型相关分析,得到二组典型变量v1‑
u1和v2‑
u2;其中:
[0016]
v1=
‑
3.387y1 0.649y2,
[0017]
u1=0.487x1 0.251x2‑
0.037x3 0.374x4‑
0.155x5 3.537x6 3.385x7‑
0.644x8 6.920x9 0.043x
10
‑
0.537x
11
;
[0018]
v2=
‑
1.181y1 3.331y2,
[0019]
u2=1.166x1 1.632x2‑
1.733x3‑
3.153x4‑
5.206x5‑
0.206x6‑
0.377x7‑
5.821x8 0.308x9 0.012x
10
0.107x
11
;
[0020]
s6.根据二组典型变量v1‑
u1和v2‑
u2得出单株毛竹扩散能力与关键调节因子的关系:
[0021]
毛竹平均扩散速率与香农
‑
维纳多样性指数、皮卢均匀度指数、多样性混交度呈负相关;
[0022]
单株中心竹株平均每年新增的毛竹株数与物种丰富度指数、玛格列夫丰富度指数、树种简单混交度呈负相关;
[0023]
所述物种丰富度指数、玛格列夫丰富度指数、香农
‑
维纳多样性指数、皮卢均匀度指数、树种简单混交度、多样性混交度作为调节单株毛竹扩散能力的关键调节因子。
[0024]
为了达到更好的发明效果,进行以下优选:
[0025]
s2步骤中,新增毛竹计算原则包括:
[0026]
(a)当两个以上样圆交集时,交集内的新增毛竹按照就近原则计算;
[0027]
(b)当两个以上样圆交集时,样圆相交点上的新增毛竹,以算术平均法计算。以提高测算准确度。
[0028]
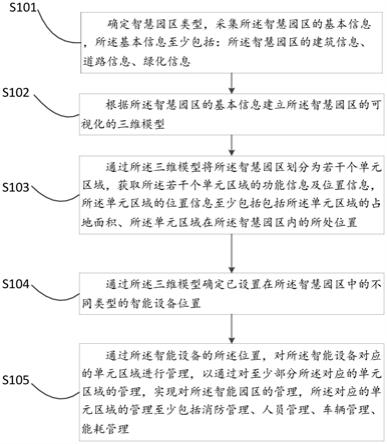
进一步优选地,s2步骤中,距样地四周边界向样地内设置10米范围作为缓冲区,以
进一步提高测算准确度。
[0029]
s2步骤中,新增毛竹计算原则还包括:(c)缓冲区内只统计新增竹株。
[0030]
所述两组典型变量v1‑
u1和v2‑
u2在表达林分结构与单株毛竹扩散能力的相关关系的贡献率和显著性水平为v1‑
u1大于v2‑
u2。
[0031]
本发明测算单株毛竹扩散能力及关键调节因子的方法可用于调节毛竹扩散能力。
[0032]
本发明中,所述林分密度、平均胸径、平均树高、物种丰富度指数、玛格列夫丰富度指数、香农
‑
维纳多样性指数、皮卢均匀度指数、树种简单混交度、多样性混交度、竞争指数和聚集指数等的计算均采用现有计算方法计算。
[0033]
本发明具有以下有益效果:
[0034]
本发明方法较现有的人工开挖地下竹鞭后人工计数单株毛竹的新增竹株数等方式计算单株毛竹扩散能力的方法更省力省时,效率明显提高,而且不会产生因开挖竹鞭产生对土壤结构、微生物的变化及影响。
具体实施方式
[0035]
以下结合具体实施例对本发明作进一步详细描述,然而,具体实施例能够以多种形式实施,且不应被理解为限于在此阐述的实施例;相反,提供这些实施例使得本发明将全面和完整,并将示例实施例的构思全面地传达给本领域的技术人员。
[0036]
本发明在浙江省磐安县黄檀林场进行了测算单株毛竹扩散能力及关键抑制因子方法研究,研究过程如下:
[0037]
1.研究目的
[0038]
研究单株毛竹扩散能力及关键抑制因子测算方法,为探明毛竹林扩张趋势及提出科学有效的抑制技术提供科学依据。
[0039]
2.研究区概况
[0040]
黄檀林场东经120
°
16
′‑
120
°
26
′
,北纬28
°
42
′‑
28
°
52
′
。属中、低山地貌,平均海拔900m。黄檀林场属中亚热带季风气候,四季分明,温暖湿润,水气光热条件良好。年平均气温为14℃至16℃,1月份平均气温
‑
0.6℃至0.9℃,7月份平均气温23℃至24.4℃,年最高气温34℃,最低气温
‑
14℃,年降雨量为1500
‑
1800mm,无霜期为185d
‑
200d。土壤类型主要为黄壤,土层深厚肥沃,理化性质较好。森林植被属于中亚热带常绿阔叶林北部亚地带,浙闽山丘甜槠木荷区。植被类型丰富,依海拔由高到低依次主要分布着山地草灌丛、针叶林、针阔混交林、毛竹林、油茶(camellia oleifera)等植被类型。黄檀林场以针叶林面积最大,主要树种有黄山松(pinus taiwanensis)、杉木(cunninghamia lanceolata)、柳杉(cryptomeria fortunei)、柏木(cupressus funebris)等;下层植被主要有杜鹃(rhododendron simsii)、茅栗(castanea seguinii)、白栎(quercus fabri)、胡枝子(lespedeza bicolor)、化香(platycarya strobilacea)、蕨科(pteridiaceae)植物等。
[0041]
3.样地调查与数据处理
[0042]
3.1样地设置与调查
[0043]
2018年10月,在黄檀林场选取毛竹纯林、竹阔混交林、分布有毛竹的杉木林、分布有毛竹的针阔混交林及分布有毛竹的阔叶林,共设置16块样地(见表1),距离样地四周边界向样地内设置10m的缓冲区,各样地规格为50m
×
50m,将样地西南角设为原点,东西向设为
横轴(x),南北向设为纵轴(y),并将各样地划分为25个10m
×
10m的小样方。对样地内所有毛竹及胸径(dbh)≥5cm的木本植物个体进行测树因子检测,主要包括:物种登记与鉴别、胸径、树高和位置坐标(x、y)(采用相邻网格法记录位置坐标),并用铁丝或钢钉套挂具有唯一编号的特制铝牌。
[0044]
表1样地基本概况
[0045][0046][0047]
3.2林分结构特征指标测定
[0048]
林分结构特征包括林分非空间结构特征与空间结构特征。
[0049]
林分非空间结构特征包括林分密度、平均胸径、平均树高(h)和物种多样性等。多样性指数以相对重要值为测算依据,采用物种丰富度指数s(即样地中的物种数)、玛格列夫丰富度指数m(margalef丰富度指数)、香农
‑
维纳多样性指数w(shannon
‑
wiener多样性指数)和皮卢均匀度指数p(pielou均匀度指数)来评价物种多样性。
[0050]
林分空间结构主要采用描述树种空间隔离程度的树种简单混交度(m1)和多样性混交度(m2)、体现林木个体所承受压力的竞争指数(ci)和反映林木个体水平分布格局的聚集指数(r)4个参数对样地林分的空间结构分析,本研究选定竞争半径为5m,相邻木4株。
[0051]
3.3毛竹扩散能力的测定
[0052]
2020年10月复查样地,在样地内分别以2018年调查时标记的单株毛竹为圆心(该毛竹记作:中心竹株),10m为半径的样圆内测定新增毛竹株数及各新增毛竹株数分别距中心竹株的距离,再根据样地调查间隔期(2年),计算单株毛竹扩散能力指标,具体公式如下:
[0053]
[0054][0055][0056]
式中,r
i
为第i株中心竹株的扩散速率,d
ij
为第i株中心竹株与第j株新增毛竹的距离,a为样地调查间隔期2年,k
i
为第i株中心竹株10米半径样圆内的新增毛竹株数,j为第i株中心竹株10米半径样圆内新增的第j株毛竹,r1为样地内n株中心竹株的平均扩散速率即毛竹平均扩散速率,r2为单株中心竹株平均每年新增的毛竹株数。
[0057]
毛竹扩散能力指标测算时,注意需要补充说明的问题如下:
[0058]
(1)当两个以上样圆交集时,交集内的新增毛竹按照就近原则计算;
[0059]
(2)当两个以上样圆交集时,样圆相交点上的新增毛竹,以算术平均法计算。
[0060]
(3)样地设置10m的缓冲区,缓冲区内只统计新增竹株,不测算2018年毛竹的扩散指标。
[0061]
3.4毛竹扩散的关键调节因子分析
[0062]
根据16个样地调查数据,以r1样地内n株中心竹株的平均扩散速率(记作:y1)、r2单株中心竹株平均每年新增的毛竹株数(记作:y2)归为一组变量组y作为毛竹扩散能力指标,将林分密度(记作:x1)、平均胸径(记作:x2)、平均树高(记作:x3)、物种丰富度指数s(记作:x4)、margalef多样性指数m(记作:x5)、shannon
‑
wiener多样性指数w(记作:x6)、pielou均匀度指数p(记作:x7)、简单混交度m1(记作:x8)、多样性混交度m2(记作:x9)、竞争指数ci(记作:x
10
)和聚集指数r(记作:x
11
)归为一组变量组x作为林分结构指标,经标准化后进行典型相关分析。
[0063]
3.5数据处理
[0064]
通过使用forsat软件,对样地林分空间结构进行计算与典型相关分析。运用spss多重比较不同类型林分毛竹扩散能力差异。
[0065]
4结果分析
[0066]
4.1毛竹扩散能力
[0067]
通过对16块样地毛竹扩散指标的测算,得出毛竹扩散指标即样地内n株中心竹株的平均扩散速率(y1)及单株中心竹株平均每年新增的毛竹株数(y2)的描述性统计信息见表2。
[0068]
表2毛竹扩散指标描述性统计信息
[0069]
指标数量最小值最大值均值标准差y1160.004.720.8561.438y2160.003.000.3410.559
[0070]
4.2不同类型林分毛竹扩散能力差异分析
[0071]
为比较分析毛竹林、竹阔混交林、杉木林、针阔混交林和阔叶林内毛竹的扩散能力,将以样地内n株中心竹株的平均扩散速率(y1)、单株中心竹株平均每年新增的毛竹株数(y2)标准化后,经单因素方差分析,y1与y2的显著性p=0.000小于显著性水平0.05,拒绝零假设,认为控制变量不同水平下各总体均值有显著差异。方差齐次性检验(见表3)后,5种类
型林分毛竹扩张指标中y1和y2的结果显著性分别为0.000和0.000,小于显著性水平0.05,方差非齐次性,因此y1和y2采用tamhane进行树种间多重比较(见表4)。
[0072]
表3方差齐次性检验
[0073]
变量levene统计量第1自由度第2自由度显著性概率py1357.6014110.000y223.1704110.000
[0074]
毛竹林、竹阔混交林、杉木林、针阔混交林及阔叶林5种类型林分的毛竹扩张速率(即平均扩散速率)和单株中心竹平均每年新增的毛竹株数多重比较结果表明(见表4),毛竹平均扩散速率在毛竹林、竹阔混交林、杉木林、针阔混交林和阔叶林5种类型林分两两比较均存在显著性差异(p<0.05),而单株中心竹平均每年新增的毛竹株数除竹阔混交林与杉木林之间不存在显著性差异外,其余类型林分两两之间均存在显著性差异。从毛竹林、竹阔混交林、杉木林、针阔混交林和阔叶林的比较结果得到毛竹平均扩散速率和单株中心竹平均每年新增的毛竹株数的大小排序均为毛竹林>竹阔混交林>杉木林>针阔混交林>阔叶林,毛竹林和竹阔混交林中分布的毛竹株数多,扩散速率快且新增株数多,而杉木林中仅有少量毛竹分布,因此平均扩散速率较慢且平均每年新增的毛竹株数较少,针阔混交林和阔叶林因其树种组成、林分结构复杂,其平均扩散速率慢,新增毛竹株数最少。
[0075]
表4多重比较结果
[0076]
[0077][0078]
4.3毛竹扩散的关键调节因子
[0079]
运用forsat中的典型相关分析,得出结果(见表5)。由于同一变量组内各典型变量之间的样本协方差为零,不同组不对应的典型变量间的样本协方差也为零,这使得分析变量组y和变量组x之间的关系转化为只需分析从两组中提取出的相对应的典型变量之间的关系。典型载荷的统计含义就是这种转化关系的反映,它作为典型相关分析的主要结果,体现出2个变量组之间的相关程度。
[0080]
表5典型变量
[0081][0082]
第一个典型变量将平均扩散速率y1分离出来,其系数为
‑
3.387,自变量组中与之相对应的解释变量主要是shannon
‑
wiener多样性指数(x6)、pielou均匀度指数(x7)和多样性混交度(x9)(相应的系数分别为3.537、3.385和6.920)。可见毛竹平均扩散速率主要受shannon
‑
wiener多样性指数、pielou均匀度指数和多样性混交度影响。毛竹平均扩散速率与shannon
‑
wiener多样性指数、pielou均匀度指数、多样性混交度均呈负相关,表明生物多样性越丰富、物种分布越均匀,混交度越大,抑制毛竹扩散的能力越强,毛竹扩散越慢;反之,则生物多样性越单一、物种分布越不均匀,混交度越小,抑制毛竹扩散的能力越弱,毛竹扩散越快。
[0083]
第二个典型变量将单株中心竹株平均每年新增的竹株数y2分离出来,其系数为3.331,自变量组中与之相对应的解释变量主要是margalef丰富度指数(x5)、物种丰富度指数(x4)和简单混交度(x8)(相应的系数分别为
‑
5.206、
‑
3.153和
‑
5.821)。可见样地内每株毛竹周围平均每年新增毛竹数量主要受物种丰富度指数、margalef丰富度指数和树种简单混
交度影响。每株毛竹周围平均每年新增的毛竹数量与margalef丰富度指数(x5)、物种丰富度指数(x4)和树种简单混交度均呈负相关;表明margalef丰富度指数、物种丰富度指数、树种简单混交度越大,单株毛竹平均每年新增的毛竹株数越少,毛竹扩散越慢;反之,margalef丰富度指数、物种丰富度指数、树种简单混交度越小,单株毛竹平均每年新增的毛竹株数越多,毛竹扩散越快。
[0084]
表5表明毛竹扩张能力与林分结构指标的2个典型变量之间均存在相关关系,典型相关系数分别是0.999和0.939,且相关性显著,表明林分结构能够清晰充分地解释相应毛竹扩散能力的变异信息。且两组典型变量(v1‑
u1)和(v2‑
u2)的方差贡献率分别为53.27%和46.73%,表明第一对典型变量(v1‑
u1)在表达林分结构与毛竹扩散能力的相关关系的贡献率及显著性水平p值高于第二对典型变量(v2‑
u2)。
[0085]
采用本发明方法测算得到的毛竹扩散能力比人工开挖地下竹鞭更省力省时,效率明显提高,而且不会产生因开挖竹鞭产生对土壤结构、微生物的变化及影响。
[0086]
本发明方法用于测算有毛竹分布的林分。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。