1.本发明属于生物技术领域,具体涉及水稻胞间连丝基因及其突变基因、编码的蛋白和应用。
背景技术:
2.水稻(orvza sativa l.)是世界上最重要的粮食作物之一,全世界有近半数的人口以稻米为主食。杂交水稻的种植,显著提高了水稻的产量,从以前的每亩200-300斤,到现在的每亩1000斤,与第二次绿色革命一起差不多解决了全球的粮食危机,为保障国家乃至世界粮食安全作出了巨大贡献。但近十几年来水稻的产量停滞不前,这一方面是由于在育种技术上没有新的突破以及遗传多样性在栽培品种中的逐步变窄,另一方面也是因为频繁发生的病虫害以及旱灾等自然灾害使得水稻生产损失惨重。然而,世界人口的持续增长以及社会经济的快速发展导致对粮食的需求不断增加。培育高产优质水稻新品种一直是分子生物学家和育种家的共同目标,而水稻籽粒数量的增加或体积的增大有利于提高水稻产量。
3.由于传统杂交育种存在育种过程周期长、杂交后代会出现性状分离及缺乏优良种质资源等问题,利用传统杂交育种技术来提高水稻产量已经无法满足人们日益增长的粮食需要。21世纪生物科学技术迅猛发展,分子生物学技术和基因工程育种技术广泛应用于实际生产,在提高水稻产量,改良水稻品质及提高水稻抗性中起到重要作用,开辟了水稻育种新时代。而结合基因工程育种技术与传统杂交育种技术来提高作物产量已成为遗传改良研究的新热点。
4.水稻籽粒增大主要是由细胞数量增加或者是由细胞体积增大引起的。胚乳占籽粒重量的80-85%,是水稻籽粒重要的组成部分,胚乳细胞的膨胀和细胞内含物质含量的增加对提高产量起着重要的作用,研究胚乳细胞体积增大的机制已成为研究热点。淀粉是水稻胚乳的主要贮藏物质,约占糙米干重的90%,其在种子中的含量直接影响水稻的产量。叶片通过光合作用合成的光合产物以蔗糖的形式通过胞间连丝汇集到维管束周围并装载到维管束韧皮部中,在韧皮部中蔗糖顺浓度梯度运输至需糖的库组织如根、幼叶、籽粒等。因此,由水稻叶片合成的碳水化合物作为合成淀粉的原料,而胞间连丝是碳水化合物运输的主要通道。故,解析胞间连丝的形成及调控机制,挖掘胞间连丝形成及调控基因,并将其应用到水稻高产新品种分子育种应用研究中,对水稻的遗传育种具有重要意义。
5.但目前尚未有此方面的研究报道。
技术实现要素:
6.有鉴于此,为了克服现有技术的不足,本发明提供一种水稻胞间连丝基因postman1及其突变基因postman1d,该基因突变后的水稻postman1-d突变体幼叶基部维管束韧皮部胞间连丝增加,且从苗期开始postman1-d突变体叶尖黄化并积累大量淀粉、蔗糖和葡萄糖,通过杂交发现该性状为显性性状,对水稻的遗传育种具有重要意义。
7.本发明提供的水稻胞间连丝基因postman1及其突变基因,所述水稻胞间连丝基因postman1位于水稻第7条染色体的标记rm20776和rm6663之间,其核苷酸序列如seq id no.14所示。
8.进一步,所述突变基因postman1d在第7条染色体loc_os07g01520基因编码框上第421碱基t-a转换,并导致第141位的编码氨基酸序列发生酪氨酸(tyrosine)到天冬酰胺(asparagine)的变异,所述突变基因核苷酸序列如seq id no.15所示。
9.进一步,所述水稻胞间连丝基因postman1编码的蛋白质,其氨基酸序列如seq id no.16所示。
10.进一步,所述水稻胞间连丝突变基因postman1d编码的蛋白质,其氨基酸序列如seq id no.17所示。
11.本发明还提供一种获得含有上述突变基因postman1d的突变体的方法,是利用甲基磺酸乙酯ems诱变缙恢10号获得遗传稳定的水稻叶片积累糖并黄化,且幼叶基部维管束韧皮部胞间连丝增加的突变体postman1-d。
12.本发明还提供一种获得上述基因及突变基因的特异引物对,其上游引物为postman1f:5
’‑
atgggtttcaatccgccggtgc-3’(seq id no.12);下游引物为postman1 r:
[0013]5’‑
ctactggtcgtggacgatgagcttg-3’(seq id no.13)。
[0014]
本发明还提供上述水稻胞间连丝突变基因postman1d在水稻高产分子育种中的应用。
[0015]
上述水稻高产分子育种中的应用,是将突变基因postman1d进行克隆并构建至植物表达载体,借助所述载体将所述基因转化至水稻体内获得过表达的植物后代。
[0016]
进一步,所述水稻为缙恢10号。
[0017]
本发明利用甲基磺酸乙酯(ems)诱变自育优良恢复系缙恢10号获得一个遗传稳定的水稻叶片积累糖并黄化,且幼叶基部维管束韧皮部胞间连丝增加的突变体postman1-d,在遗传分析和基因定位的基础上,先通过基因预测、同源搜索及基因序列差异比较,初步确定了水稻突变性状为postman1显性基因控制,postman1编码未知功能蛋白(loc_os07g01520)。随后,本发明以该显性突变体postman1-d为材料,克隆了水稻基因postman1,具有如seq id no.14所示的核苷酸序列,开放阅读框为2445bp,由1个外显子,无内含子组成,编码814个氨基酸,其氨基酸序列如seq id no.16所示。与野生型缙恢10号相比,突变基因postman1d在第7染色体loc_os07g01520基因编码框上第421碱基有t-a的转换,并导致第141位的编码氨基酸序列发生酪氨酸(tyrosine)到天冬酰胺(asparagine)的变异,其核苷酸序列如seq id no.15所示,其编码氨基酸序列如seq id no.17所示。
[0018]
然后,构建功能互补载体并转化野生型缙恢10愈伤组织。经鉴定转基因阳性植株叶片黄化,与显性postman1-d突变体表型一致。进一步确定水稻叶片积累糖并黄化,且幼叶基部维管束韧皮部胞间连丝增加的突变性状由postman1基因突变引起。
[0019]
本发明的有益效果在于:本发明提供了水稻胞间连丝基因postman1及突变基因postman1d,该基因调控胞间连丝发育,促进碳源分配,对水稻的遗传育种具有重要意义。
附图说明
[0020]
为了使本发明的目的、技术方案和有益效果更加清楚,本发明提供如下附图:
[0021]
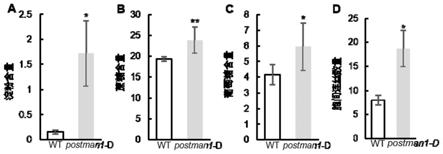
图1为野生型缙恢10号(wt)与水稻胞间连丝突变体postman1-d中糖含量测定结果及幼叶维管束韧皮部胞间连丝数量统计结果(其中a为淀粉含量;b为蔗糖含量;c为葡萄糖含量;d为胞间连丝统计结果)。
[0022]
图2为水稻胞间连丝基因postman1基因的遗传和物理图谱(a为postman1的初定位区间在第7染色体长臂ssr标记rm20776和rm6663间;b为将postman1基因精细定位在标记idel7-9与idel7-14间37kb的范围内;c为定位区间预测基因;d为突变体postman1的候选基因loc_os07g01520的结构及突变位置)。
[0023]
图3为postman1基因编码蛋白的进化树分析结果。
[0024]
图4为互补重组载体构建图。
[0025]
图5为postman1突变体互补表型分析,其中wt为野生型,postman1-d为显性胞间连丝突变体,postman1-d::com为postman1-d突变基因转化野生型缙恢10(wt)愈伤组织的转基因阳性植株。
具体实施方式
[0026]
以下将参照附图,对本发明的优选实施例进行详细的描述。优选实施例中未注明具体条件的实验方法,通常按照常规条件,例如分子克隆实验指南(第三版,j.萨姆布鲁克等著,黄培堂等译,科学出版社,2002年)中所述的条件,或按照制造厂商所建议的条件。
[0027]
本发明实施例中使用的材料:野生型水稻材料缙恢10号(wt)和显性胞间连丝突变体(postman1-d),均由本实验室培育;m-mlv逆转录酶、高保真dna聚合酶pfu、taq dna聚合酶、t4 dna连接酶、限制性内切酶、pmd19-t载体、trizol试剂盒、dna凝胶回收试剂盒、质粒提取试剂盒、dna marker购自takara公司;dna marker iii购自天根生化科技(北京)有限公司;氨苄青霉素(ampicillin,amp)和卡拉霉素(kanamycin,kan)为sigma公司产品;引物合成和dna测序由擎科生物有限公司完成;其它化学试剂购自北京鼎国生物技术有限责任公司;大肠杆菌dh5α、农杆菌eha105由本实验室保存。
[0028]
实施例1、水稻显性胞间连丝突变体postman1-d的获得和形态学观察
[0029]
利用甲基磺酸乙酯(ems)诱变自育优良恢复缙恢10号获得一个遗传稳定的水稻叶片积累糖并黄化,且幼叶基部维管束韧皮部胞间连丝增加的突变体,命名为postman1-d。水稻突变体postman1-d的叶尖从苗期开始表现出黄化表型,该表现持续到成熟期,该突变体叶片中积累了大量的糖包括淀粉、葡萄糖、蔗糖,且该突变体幼叶基部维管束韧皮部胞间连丝数量增加(图1a-d)。表明postman1-d可能是通过调控胞间连丝形成从而影响水稻叶片糖分配,进而影响叶片功能导致叶片黄化。该性状经过多代观察,表现稳定遗传。
[0030]
实施例2、胞间连丝postman1基因遗传分析与定位
[0031]
以postman1-d突变体为父本,籼稻品种西农1b(xinong1b)为母本杂交获得f1代植株叶片全部表现为叶尖黄化,然后通过自交获得22889株f2代群体中,依据黄叶性状分离出了突变叶片和正常叶片两种表型,分离出7424株正常株,其余为突变株,可以看出正常株与突变株符合1:3的分离比例,表明该突变性状由一对显性单基因控制。
[0032]
初步定位:选取均匀分布于水稻12条染色体上的480对ssr引物,在亲本postman1-d和西农1b间检测多态性,其中有98对ssr引物显示多态性。用这98对引物在正常和突变基因池中进行基因连锁分析,筛选与基因postman1连锁的ssr标记,发现postman1与第7染色
体短臂上标记rm20776和rm6663连锁。用连锁标记rm20776和rm6663分析200株隐性定位群体,结果显示,基因postman1位于rm20776和rm6663之间(图2a)。
[0033]
精细定位:根据已公布的籼稻品种93-11序列,在标记rm20776和rm6663间进一步筛选和开发了ssr引物及indel引物,其中有3对在亲本间表现多态性(表1)。用3对具有多态性的ssr标记及2对idel标记序列分析所有7224株突变单株,结果表明:标记rm20808、idel7-9和idel7-14与基因postman1之间的遗传距离分别为0.048cm、0.007cm、和0.007cm(图2b)。最终postman1被定位在标记idel7-9和idel7-14之间约37kb的范围内(图2c)。对该定位区间进行分析(http://www.gramene.org),发现该定位区间内包含6个编码基因(图2c)。
[0034]
表1、3对具有多态性的ssr标记及2对idel标记序列
[0035]
引物正向序列(5
′→3′
)反向序列(5
′→3′
)大小(bp)brm20776cctcatgttccctttccggttgg(seq id no.1)taagggcacagtaggcgggaac(seq id no.2)96rm6663acaaatacagtggaagcgtgtcg(seq id no.3)gaacacgtctgggagcactacg(seq id no.4)133rm20808ggggaatactccatttgtacaagc(seq id no.5)gaactcaatcacacatggaacgc(seq id no.6)156idel7-9cacatagcaccagttaatttacctc(seq id no.7)cggttggtgttattaaccggg(seq id no.8)164idel7-14gcggatagtccggatacgg(seq id no.9)gctaggttgaaggtctagagc(seq id no.10)177
[0036]
实施例3、克隆loc_os07g01520基因
[0037]
根据genbank已登录的水稻日本晴基因loc_os07g01520序列(loc_os07g01520基因cdna(seq id no.11)),利用vector nti软件设计扩增postman1-d突变体和野生型缙恢10号loc_os07g01520序列的特异引物:上游引物postman1 f:5
’‑
atgggtttcaatccgccggtgc-3’(seq id no.12);下游引物postman1 r:5
’‑
ctactggtcgtggacgatgagcttg-3’(seq id no.13)。
[0038]
分别取野生型缙恢10号和突变体postman1-d在光照培养两周的幼叶2g,迅速放入液氮中研磨成粉末,按照trizol试剂盒说明书提取总rna。所得野生型缙恢10号和突变体postman1-d总rna的电泳结果显示主带清晰完整,28s和18s的条带亮度比约为2:1,说明rna的浓度和纯度符合实验要求,可以用于合成双链cdna。然后分别以所得野生型缙恢10号和突变体postman1-d总rna为模板,按照m-mlv逆转录酶说明书,使用oligo(dt)引物进行逆转录获得cdna;再以cdna为模板,以seq id no.12和seq id no.13所示序列为特异引物及高保真dna聚合酶pfu进行pcr扩增,pcr反应条件为:94℃预变性5分钟;然后94℃变性30秒,55℃复性30秒,72℃延伸1分钟,共35个循环;最后72℃延伸10分钟。将rt-pcr产物进行1.0%(g/ml)琼脂糖凝胶电泳检测。结果显示,野生型缙恢10号和突变体postman1-d的扩增产物均在约2400bp处呈单一特异性条带,并将野生型缙恢10号扩增产物命名为postman1基因,突变体postman1-d扩增产物命名为postman1突变基因(postman1d)。
[0039]
然后按照dna凝胶回收试剂盒说明书进行切胶回收纯化,纯化的postman1基因和postman1d突变基因与pmd-19t载体在t4 dna连接酶的作用下于16℃连接过夜,连接产物转化大肠杆菌dh5α感受态细胞,用含有氨苄青霉素的lb平板筛选阳性克隆,提取质粒,pcr鉴定后测序,分别得克隆载体pmd-19t-postman1和pmd-19t-postman1d。将克隆载体pmd-19t-postman1和pmd-19t-postman1d送测序公司进行测序,结果显示野生型postman1基因序列如seq id no.14所示,无内含子,开放阅读框为2445bp。突变基因postman1d序列如seq id no.15所示,开放阅读框为2445bp,与野生型缙恢10号相比,突变基因postman1d在编码框第
424碱基有t-a的转换,并导致第141位的编码氨基酸序列发生酪氨酸(y)到天冬酰胺(n)的变异,突变后氨基酸序列如seq id no.17所示。
[0040]
将获得的水稻胞间连丝基因postman1编码蛋白的氨基酸序列进行结构域分析,发现该基因编码蛋白具有两个未知功能的结构域duf4220和duf594,结果如图2d所示。
[0041]
实施例4、突变基因postman1的生物信息学分析
[0042]
从ncbi及phytozome查找水稻(oryza sativa)loc_os07g01840、loc_os10g20770,玉米(zea mays)zm00008a10535,谷子(setaria italica)seita.j002000、seita.j001900,狗尾巴草(setaria viridis)sevir.2g000400、sevir.2g000500、sevir.2g000600,高粱(sorghum bicolor)sobic.0002g004200、sobic.0002g004300、sobic.0002g004400,(brachypodium stacei)brast06g242900、brast06g243000,二穗短柄草(brachypodium distachyon)bradi1g59470、bradi1g59480、bradi2g21170,菠萝(ananas comosus)aco021351、aco031804、aco014422、aco018136,苜蓿(medicago truncatula)medtr1g090683、medtr1g090687,巨桉(eucalyptus grandis)eucgr.b03804、eucgr.b03807、eucgr.j01172,雷蒙德氏棉(gossypium raimondii)gorai.004g181100、gorai.012g050500、gorai.008g221400、gorai.004g181100,gorai.008g221400,小果野芭蕉(musa acuminata)gsmua_achr6t21850,洛矶山耧斗菜(aquilegia coerulea)aqcoe4g263800、aqcoe4g251900、aqcoe1g235800、aqcoe4g300100,aqcoe4g290400,拟南芥(arabidopsis thaliana)at5g45540、at5g45530,白菜型油菜(brassica rapa)brara.f03680、brara.f03681氨基酸序列。保守结构域分析(smart:)发现以上蛋白均编码未知功能蛋白且都具有两个未知功能保守结构域duf4220和duf594。利用megx软件进行氨基酸序列的比对及进化树的生成,结果如图3所示。结果显示,突变蛋白postman1在水稻中有两个同源蛋白loc_os07g01840、loc_os10g20770,同源性分别为75%、60%,此外,与高粱sobic.0002g004200(59%)和二穗短柄草bradi1g59480(61%)的亲缘性较近。且突变基因postman1的突变位点发生在duf4220结构域内。
[0043]
实施例4、突变基因postman1的功能验证
[0044]
为了验证水稻突变体postman1-d的突变性状是由突变基因postman1引起的,扩增突变体postman1-d中loc_os07g01520基因组片段包括2432bp 5’端非编码区,2445bp全长编码框,364bp 3’非编码区通过ecorⅰ和hind iii连入pcambia1301载体中获得重组表达载体pcambia1305-postman1d::com,其结构如图4所示。将获得的重组表达载体pcambia1305-postman1d::com转化野生型缙恢10号,获得转基因植株,然后观察转基因植株的性状,结果如图5所示。结果显示,转基因植株叶尖黄化且积累大量淀粉,表型与postman1-d突变体相似,进一步证实了postman1-d突变体是由loc_os07g01520基因编码框第424位碱基由“t”突变为“a”引起的。
[0045]
淀粉是水稻胚乳的主要贮藏物质,约占糙米干重的90%,其在种子中的含量直接影响水稻的产量。由水稻叶片合成的碳水化合物是合成淀粉的前体,而胞间连丝是碳水化合物运输分配的主要通道,促进胞间连丝的发育可以促进水稻碳源的分配运输,从而促进水稻植株的生长发育及籽粒灌浆过程。解析胞间连丝的形成及调控机制,挖掘胞间连丝形成及调控基因,并将其应用到水稻高产新品种分子育种应用研究中具有重要的意义。故,胞间连丝调控基因postman1及其突变基因postman1d可作为高产优质水稻新品质分子育种研
究的候选基因。
[0046]
最后说明的是,以上优选实施例仅用以说明本发明的技术方案而非限制,尽管通过上述优选实施例已经对本发明进行了详细的描述,但本领域技术人员应当理解,可以在形式上和细节上对其作出各种各样的改变,而不偏离本发明权利要求书所限定的范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。