氨基酸转运蛋白csaap7.2及其编码基因和应用
技术领域
1.本发明涉及茶树分子遗传育种领域,具体涉及氨基酸转运蛋白csaap7.2及其编码基因和应用。
背景技术:
2.茶叶质量在很大程度上取决于茶加工用新芽中多酚、咖啡因和茶氨酸的含量。这些代谢产物的生物合成与氮(n)条件有关,高质量的茶叶的生产需要提供适当的n水平。n水平过高或者过低均会对茶叶质量产生较大影响。例如,缺n使得茶树植株发育不良,植株矮小,高施氮肥会加速土壤酸化,导致茶叶中铝、氟和重金属(如铅和铬)大量积累,从而对人类健康造成潜在风险。近几十年来,有机种植园生产的茶叶越来越受欢迎。与过去二十年相比,有机茶产量增长了45倍以上。
3.此前,研究表明,植物以硝酸盐、铵、尿素和氨基酸的形式从土壤中获取氮(williams &miller,2001),而氨基酸是某些土壤中重要的氮库(friedel&scheller,2002;yu et al., 2002;kranabetter et al.,2007)。土壤氨基酸主要来源于腐朽生物的蛋白质和肽的外酶分解。茶树是多年生落叶作物,每年需要定期修剪2~3次,以保持旺盛的营养生长。通常,茶园每年产生约8000kg/ha的修剪凋落物。这种修剪过的枯枝落叶含有大量的氨基酸和蛋白质,在土壤中分解后可以循环利用。这些修剪凋落物衍生的氨基酸可能是茶树根系吸收的重要氮源。
4.植物细胞,包括根细胞,通过被动和主动运输机制相结合的方式吸收养分。当土壤养分浓度较高时,通道和渗透可以参与被动吸收,而质子耦合转运蛋白在低养分条件下参与次级主动转运步骤。鉴于土壤氨基酸水平远低于根细胞内的氨基酸水平,通常需要质膜转运蛋白从土壤中吸收氨基酸。目前,许多植物氨基酸转运蛋白已被鉴定,并被分为两个超家族: aaap(氨基酸/生长素渗透酶)和apc(氨基酸多胺胆碱转运蛋白)。aaap超家族包括六个家族:aap(氨基酸渗透酶)、lht(赖氨酸和组氨酸转运体)、prot(脯氨酸转运体)、 gat(γ-氨基丁酸转运体)、aux(生长素转运体)和ant(芳香族和中性氨基酸转运体),而apc超家族包括cat(阳离子氨基酸转运体)和lat(l型氨基酸转运体)家族。影响土壤吸收的氨基酸转运蛋白主要属于aap、prot和lht家族。我们前期已经鉴定到6 个茶树aap家族能够转运氨基酸的氨基酸转运蛋白,即csaap1、csaap2、csaap4、 csaap5、csaap6和csaap8。但是是否有aap家族基因提高茶树根系中的氨基酸含量,进一步提高氨基酸的地下向地上组织运输并不清楚。因此,深入研究茶树根系吸收氨基酸的分子机制,将为通过基因工程手段增加其从土壤中吸收氨基酸、提高氮素利用率,进而提高茶树新稍中的氨基酸含量。
技术实现要素:
5.针对现有技术的不足,本发明的目的在于提供一种氨基酸转运蛋白csaap7.2及其编码基因和应用。
6.本发明的目的可以通过以下技术方案实现:
7.第一方面,本发明保护一种调控茶树根系吸收转运氨基酸的转运蛋白csaap7.2,所述蛋白为下述任一种:
8.(1)其氨基酸序列如seq id no.2所示;
9.(2)将seq id no.2所示的氨基酸序列经过氨基酸残基的取代和/或缺失和/或添加得到的与(1)所示的蛋白质具有80%同一性且具有相同功能的蛋白质。
10.第二方面,本发明还保护编码前文所述的转运蛋白csaap7.2的基因。
11.作为本技术的优选技术方案,所述基因具有如下a1)或a2):
12.a1)其核苷酸序列如seq id no.1所示;
13.a2)与seq id no.1所示的序列具有至少80%、至少85%、至少90%、至少95%、至少 98%或至少99同源性的核苷酸序列。
14.本发明还保护扩增前文所述基因的开放阅读框序列的引物,其核苷酸序列如seq idno.3/seq id no.4所示。
15.本发明还保护含有前文所述基因的表达盒、表达载体、转基因细胞系或重组工程菌。
16.优选的,所述表达载体为亚细胞定位载体、酵母表达载体和拟南芥过表达载体。
17.第三方面,本发明保护一种提高植物根系从外源吸收转运氨基酸能力的方法,所述方法包括提高目的植物中前文所述的蛋白的含量和/或活性。
18.作为本技术的优选技术方案,所述提高目的植物中前文所述的蛋白的含量和/或活性通过提高目的植物中所述蛋白的编码基因的表达量实现。
19.作为本技术的优选技术方案,所述提高目前植物中所述蛋白的编码基因的表达量通过将前文所述蛋白的编码基因导入目的植物来实现。
20.优选的,所述目的植物为拟南芥或茶树。
21.第四方面,本发明还保护前文所述的蛋白,和/或,前文所述的基因,和/或,前文所述的引物,和/或,前文所述的含有前文所述基因的表达盒、表达载体、转基因细胞系或重组工程菌的下述任一种应用:
22.b1)在调节植物根系从外源吸收转运氨基酸中的应用;
23.b2)在制备调节植物根系从外源吸收转运氨基酸的产品中的应用;
24.b3)在培育高氮素利用率的植物中的应用;
25.b4)在植物育种中的应用。
26.作为本技术的优选技术方案,所述植物为拟南芥或茶树。
27.本发明的有益效果
28.本发明构建了茶树csaap7.2基因的酵母表达载体,并将其转化到氨基酸吸收缺陷型的酵母突变体菌株22δ10α中,获得的重组菌株经功能分析,表明其具有氨基酸转运活性。
29.本发明还构建了csaap7.2基因的植物过表达载体并转化到野生型拟南芥中,获得的转基因植株经生物学功能验证,表明其具有介导植物根系从外源吸收氨基酸的作用。
30.本发明提供的csaap7.2介导茶树根系从外源吸收氨基酸的功能,为生产上施用氨基酸肥提供理论依据,为培育高氮素利用率的茶树新种资提供基因资源。
附图说明
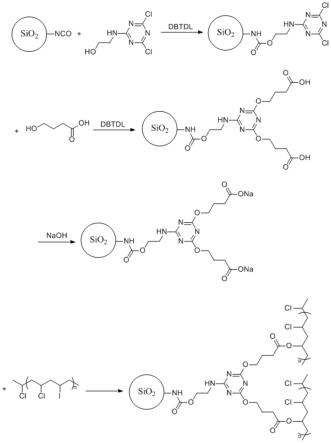
31.图1是csaap7.2的组织特异性表达图;
32.图2是csaap7.2的亚细胞定位图;
33.图3是csaap7.2在酵母中的功能鉴定图;
34.图4是csaap7.2在拟南芥中的过量表达图。
具体实施方式
35.以下为对本发明具体实施例做出的具体描述,用来详细说明本发明,但并不以任何方式限制本发明的应用范围。在不偏离本发明基本特征的前提下,本领域技术人员可以对本发明做出任何形式的改变和修改,以使其能够更便于被应用。下述实施例中的实验方法和试验材料,如无特殊说明,均为常规方法和常规生化试剂。
36.申请人从茶树中分离克隆得到一个新的氨基酸转运蛋白基因csaap7.2,其核苷酸序列如seq id no.1所示,cds全长序列为1395bp;该基因编码465个氨基酸,等电点为 4.93,分子量为51.4kda,氨基酸序列如seq id no.2所示。
37.本发明还提供了用于扩增所述csaap7.2基因开放阅读框的引物,其核苷酸序列如seq id no.3/seq id no.4所示;用于qrt-pcr技术分析的荧光定量引物,其核苷酸序列如seq id no.5/seq id no.6,所用内参基因为csgapdh,其核苷酸序列如seq id no.7/seq id no.8所示;用于构建氨基酸转运蛋白csaap7.2基因的亚细胞定位载体、酵母表达载体和拟南芥过表达载体引物,其核苷酸序列分别如seq id no.9/seq id no.10,seq id no.11/seq id no.12和seq id no.13/seq id no.14所示;
38.本发明还提供了含有所述氨基酸转运蛋白基因csaap7.2的亚细胞定位载体、酵母表达载体和拟南芥过表达载体,及对应的重组菌株。
39.本发明还提供了所述基因csaap7.2、所述蛋白csaap7.2,在调节植物根系从外源吸收转运氨基酸中的应用。
40.本发明构建茶树氨基酸转运蛋白基因csaap7.2的酵母表达载体并转化氨基酸吸收缺陷型的酵母突变体菌株22δ10α中,获得的重组菌株在氨基酸为唯一氮源培养基上的生长缺陷表型得到部分恢复。
41.本发明构建的基因csaap7.2的植物过表达载体并转化到野生型拟南芥中,以转化空载体的拟南芥植株为对照,获得csaap7.2基因表达量明显高于对照的拟南芥过表达株系,该过表达植株表现为对外源茶氨酸的吸收能力增强,和对高浓度茶氨酸不耐受的表型。
42.实施例1,茶树csaap7.2基因全长的克隆
43.以一年生
‘
苏茶早’茶树的根为试验材料,参照植物多糖多酚总rna提取试剂盒(购自天根公司)的说明书提取总rna,参照ii one step rt-pcr kit with gdna eraser 试剂盒(购自诺唯赞公司)的说明书进行cdna第一条链的合成。
44.参考ncbi数据库提供的基因序列,使用primer premier 5.0软件设计基因全长克隆引物。利用上述合成的cdna为模板通过pcr技术用于csaap7.2基因的全长扩增,其引物对的核苷酸序列如下所示:
45.f1:5
’‑
atgggggatgaaggagaaga-3’(seq id no.3);
46.r1:5
’‑
ttagctccattttgcagctata-3’(seq id no.4)。
47.pcr反应体系和程序参考max dna polymerase说明书。将获得的pcr产物经1%的琼脂糖凝胶电泳,获到单一目的条带的pcr产物,按照胶回收试剂盒(购自诺唯赞公司)使用说明提取步骤,回收该目的条带。
48.实施例2,茶树不同组织中氨基酸转运蛋白csaap7.2基因的qpcr分析
49.以一年生
‘
苏茶早’茶树的根、茎、第1叶、第3叶、第5叶和花为试验材料,利用实施例1中的实验方法获得cdna模板。根据实施例1扩增获得的基因全长序列,使用primer premier 5.0软件设计csaap7.2基因特异的荧光定量引物,引物的核苷酸序列如下:
50.f2:5
’‑
aagttttgggggtcttgggg-3’(seq id no.5),
51.r2:5
’‑
tcaactgacccaacgaaggc-3’(seq id no.6)。
52.本实验采用的内参基因为csgapdh,其引物对的核苷酸序列如下所示:
53.f3:5
’‑
ttggcatcgttgagggtct-3’(seq id no.7),
54.r3:5
’‑
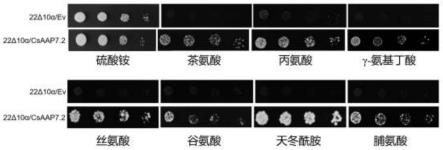
cagtgggaacacggaaagc-3’(seq id no.8)
55.qrt-pcr采用sybr green i dye试剂盒(购自诺唯赞公司),反应体系和程序参考说明书进行。
56.以茶树不同组织为材料,组织定位实验结果表明csaap7.2基因在根和茎中表达量较高,而在其他组织中的表达量较低(图1)。
57.实施例3,茶树氨基酸转运蛋白csaap7.2的亚细胞定位
58.将实例1中克隆得到的产物作为模板,再次通过带同源臂和酶切位点的引物进行扩增,引物的核苷酸序列如下:
59.f4:5
’‑
ggggacaagtttgtacaaaaaagcaggcttcatgggggatgaaggagaaga-3
’ꢀ
(seq id no.9);
60.r4:5
’‑
ggggaccactttgtacaagaaagctgggtcttagctccattttgcagctata-3
’ꢀ
(seq id no.10)。
61.利用bp克隆酶(购自英骏公司)将其连接到入门载体pdonr207上,通过菌落pcr 的方法初步挑选阳性克隆,而后将该克隆送通用公司进行测序。测序正确的质粒进行下一步实验。而后利用lr克隆酶(购自英骏公司)将其连接到终载体pk7wgf上。接着,利用热激法将重组载体pk7wgf-csaap7.2和对照空载分别转入农杆菌eha105中。表达已知内质网定位基因atwak2的农杆菌株作为阳性对照。用叶片渗透法将上述菌株注射到烟草表皮细胞中,具体操作方法:
62.1)将上述农杆菌分别在相应抗性的lb液体培养基中培养过夜;
63.2)6000g离心5分钟收集菌体;
64.3)将菌体重悬在ph为6.0的诱导液(10mm mgcl2,10mm mes,100μm acetosyringone)中,使其菌液浓度保持在od600=0.8;
65.4)室温避光2-3小时;
66.5)利用1ml注射器,将菌液注射到生长健壮的本生烟草叶片背面;
67.6)正常培养2-3天,通过激光共聚焦显微镜观察。
68.亚细胞定位结果如图2所示,注射含有pk7wgf空载质粒农杆菌的烟草表皮细胞,发现在细胞的各细胞器中均能观察到绿色荧光。而注射含有pk7wgf-csaap7.2质粒农杆菌的烟草表皮细胞,仅在细胞的部分区域观察到绿色荧光,且该绿色荧光与表达已知内质网定
位基因atwak2的红色荧光信号重叠,表明csaap7.2是一个内质网定位蛋白。
69.实施例4,茶树氨基酸转运蛋白csaap7.2的酵母异源表达
70.将实施例1中克隆得到的产物作为模板,再次通过带同源臂和酶切位点的引物进行扩增,引物的核苷酸序列如下:
71.f5:5
’‑
aagcttggtaccgagctcggatccatgggggatgaaggagaaga-3’(seq id no.11),
72.r5:5
’‑
ccctctagatgcatgctcgagttagctccattttgcagctata-3’(seq idno.12)。
73.利用限制性内切酶hind iii和bamh i,将酵母表达载体pyes2的质粒进行线性化处理。将pcr克隆得到的目的片段和线性化后的质粒,利用iione step cloningkit(购自诺唯赞公司)试剂盒进行重组连接,方法参考说明书。将构建完成质粒转化大肠杆菌感受态trans-t1,通过菌落pcr的方法初步挑选阳性克隆,而后将该克隆送通用公司进行测序。
74.将测序正确的克隆进行质粒提取,并将其转化到22δ10α酵母感受态细胞中,转化空载体作为对照。感受态制备和转化方法参考frozen-ez yeast transformationⅱkit(购自zymoresearch公司)试剂盒说明书。而后将转化后的酵母菌液涂布到缺少尿嘧啶的ynb培养基上,30℃培养3天后,挑选阳性克隆进行扩大培养。
75.将扩大培养好的菌液12000rpm离心2分钟,取上清。向菌体中加入无菌水清洗菌体3 次。将菌体悬浮在无菌水中,并将其依次稀释10-1
,10-2
和10-3
倍。并将原液和稀释后菌液依次吸取2μl到含有2mm的硫酸铵、丙氨酸、γ-氨基丁酸、丝氨酸、谷氨酸、天冬酰胺、脯氨酸或茶氨酸为唯一氮源的ynb平板上,30℃培养3天并观察生长情况。
76.结果如图3所示,表达csaap7.2基因的酵母菌株能够恢复突变体在氨基酸为唯一氮源上不能正常生长的表型,表明该基因具有氨基酸转运功能。
77.实施例5,过量表达csaap7.2基因能够增强拟南芥对外源茶氨酸的吸收
78.将实例1中克隆得到的产物作为模板,再次通过带同源臂和酶切位点的引物进行扩增,引物的核苷酸序列如下:
79.f6:5
’‑
gactctagaggatccatgggggatgaaggagaaga-3’(seq id no.13),
80.r6:5
’‑
gggaaattcgagctcttagctccattttgcagctata-3’(seq id no.14)。
81.利用限制性内切酶bamh i和sac i,将表达载体pbi121的质粒进行线性化处理。将 pcr克隆得到的目的片段和线性化后的质粒,利用iione step cloning kit(购自诺唯赞公司)试剂盒进行重组连接,方法参考说明书。将构建完成质粒转化大肠杆菌感受态trans-t1,通过菌落pcr的方法初步挑选阳性克隆,而后将该克隆送通用公司进行测序。
82.将测序正确的克隆进行质粒提取,而后利用热激法将pbi121-csaap7.2载体转化至农杆菌感受态gv3101,利用浸花法构建拟南芥过表达株系。通过喷施basta进行过表达阳性株系的筛选。对筛选获得的阳性苗,通过实施例2描述的荧光定量的方法很引物进行株系鉴定。
83.鉴定完成的3个过表达拟南芥株系种子灭菌,而后将其在含有20mm茶氨酸的1/2ms 培养基上进行培养,生长在不含茶氨酸的1/2ms培养基上作为对照,10天后进行表型观察。
84.结果如图4所示,在正常培养条件下,表达csaap7.2基因的拟南芥植株与野生型没有差异;在高浓度茶氨酸的条件下进行培养,表达csaap7.2基因的拟南芥植株较野生型拥
有更高的致死率。这一结果表明,过量表达csaap7.2基因能够增强拟南芥对外源茶氨酸的吸收功能。
85.由上述实施例可知,本发明提供的氨基酸转运蛋白基因csaap7.2,经生物学功能验证具有促进植物根系从外源吸收氨基酸的作用。
86.以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。