源自噬菌体的纳米孔传感器的制作方法
- 国知局
- 2024-07-27 12:31:04
相关申请的交叉引用
本申请要求2018年2月9日提交的美国临时申请62/628,562的权益,其全部内容通过引用并入本文。
关于联邦政府赞助的研究或开发的声明
本发明是在美国国立卫生研究院授予的eb012135号政府资助下完成的。政府对本发明具有一定的权利。
背景技术:
多种蛋白质抗原已被证明可作为癌症生物标记物,用于早期检测、诊断或预测,例如癌抗原125、人附睾分泌蛋白4、psa、前列腺特异性膜抗原(psma)等。传统的生物化学检测方法,如elisa、western印迹、质谱和荧光成像已用于诊断癌症。但是,由于这些经典方法耗时且费力的样品制备、低灵敏度或低准确性,因此在工业环境中的适用性有限。
纳米孔技术的进步激发了一系列生物医学应用,例如核酸、化学品、肽或蛋白质传感。在单分子水平上基于纳米孔的传感有望克服传统方法的一些缺点。生物和合成来源的纳米孔均用于通过可调电阻脉冲传感来特异性地识别分析物,从而使感兴趣的分析物被电动势驱动通过具有纳米尺寸的孔,并通过离子特征曲线进行识别。与合成纳米孔相反,生物纳米孔是同质的,并且易于大规模获得和纯化。晶体结构信息的可用性允许针对特定传感应用精确修饰生物纳米孔。纳米孔基于天然存在的膜蛋白,并且可以通过使膜与纯化的蛋白接触并将电压电势施加到膜上而插入到共聚物膜中。
噬菌体phi29纳米通道,其不是天然膜蛋白,已发展成为功能强大的基于纳米孔的平台,可在单分子水平上对化学品、dna、rna、肽和抗体进行指纹分析。将纳米通道插入脂质体中并与平面脂质双层融合。电动势使分析物通过phi29纳米通道,从而暂时阻止离子流并产生电子阻止信号。此外,与依附在或通过基因工程改造到phi29纳米通道的配体结合的分析物会导致在电子信号曲线中观察到通道的构象变化。然而,需要高通量方法以便在测试组中实现蛋白质测序或多种生物标记物检测。
技术实现要素:
本文公开了将噬菌体dna包装马达的连接子蛋白通道插入共聚物膜内用作纳米孔的方法。噬菌体phi29纳米通道不是天然膜蛋白,并且纯化的蛋白不能插入共聚物膜中。通过将插入连接子蛋白通道的脂质体与共聚物膜融合,实现插入。如此制备的膜是包含连接子蛋白通道的共聚物膜。共聚物膜也可以包含来自与共聚物膜融合的脂质体的脂质。
因此,还公开了包含共聚物膜的组合物,其中所述共聚物膜中已经插入了噬菌体包装马达的连接子蛋白通道。所述组合物可作为纳米孔传感器,用于生物医学应用,例如高通量蛋白质测序或癌症诊断。
将连接子蛋白通道嵌入共聚物膜中,使得它们可以用作高通量仪器中的纳米孔。例如,oxfordnanoporetechnologiesltd.(ont)已经开发了基于电子技术的高通量纳米孔传感技术,以分析各种类型的分析物。用户可以选择不同类型的ont仪器(例如:smidgion,minion,gridion,promethion)用于不同的实验,并且它们可以与不同类型的流动池配合,在所述流动池中纳米孔嵌入在共聚物膜中。ont聚合物膜至少稳定数月,并且还耐较高电压。纳米孔的这种使用允许高通量、同时检测许多类型的癌症生物标记物,并且使得能够进行诸如蛋白质测序的应用。每个通道可以包含其自己的电极对,从而分离通道之间的电信号。本文公开了将噬菌体phi29的门户蛋白插入共聚物膜的方法。在一个实施方案中,膜可以存在于适于高通量测序的装置中。
特别地,oxfordnanoporetechnologiesltd.已经开发了具有512个通道的多路复用芯片的minion装置。这种高通量检测允许同时检测许多类型的癌症生物标记物,并使得能够进行诸如蛋白质测序的应用。每个通道包含其自己的电极对,从而分离通道之间的电信号。此外,minion使用三嵌段共聚物膜,其比传统的脂质膜更稳定。本文公开了将噬菌体phi29的门户蛋白插入minion装置以用于生物医学应用如高通量蛋白质测序或癌症诊断的方法。
在一些实施方案中,将噬菌体phi29的门户蛋白插入共聚物膜的方法包括将噬菌体dna包装马达的连接子蛋白通道引入脂质体,使共聚物膜与脂质体接触,并通过跨共聚物膜施加电压诱导脂质体-聚合物融合。因此,公开了将源自噬菌体dna包装马达的连接子蛋白的纳米孔插入共聚物膜中的方法,所述方法包括使所述共聚物膜与包含所述纳米孔的脂质体接触,并且跨所述共聚物膜施加电压以诱导脂质体-共聚物融合。源自噬菌体dna包装马达的连接子蛋白的纳米孔可以是本文定义的任何纳米孔。
含有适合用于所公开的组合物和方法的连接子蛋白通道的噬菌体dna包装马达的实例包括但不限于phi29(登记号yp_002004539.1)、t3(登记号np_523332.1)、t4(登记号np_049782.1)、t5(登记号aas77191.1)、t7(登记号np_041995.1)、spp1(登记号caa66580.1)、hk97(登记号np_037699.1)、g20c(登记号kx987127.1)、噬菌体2(登记号np_046757)、噬菌体3(nutter等,1972jviral.10(3):560-2)、噬菌体22(登记号aaa72961)和lambda噬菌体(登记号gi549295、gi6723246、gi15837315、gi16764273)dna包装马达。在一些实施方案中,连接子蛋白包括噬菌体phi29连接子蛋白gp10。在一些实施方案中,连接子蛋白包括噬菌体t3连接子蛋白gp8。在一些实施方案中,连接子蛋白包括噬菌体t7连接子蛋白gp8。在一些实施方案中,连接子蛋白包括噬菌体t4连接子蛋白gp20。在一些实施方案中,连接子蛋白包括噬菌体t5连接子蛋白gp7。在一些实施方案中,连接子蛋白包括噬菌体spp1连接子蛋白gp6。在一些实施方案中,连接子蛋白包括噬菌体hk97连接子蛋白gp3。
因此,本文公开了纳米孔传感器,其包含整合到共聚物膜中的噬菌体dna包装马达的连接子蛋白通道。具体地,本文公开了包含纳米孔的共聚物膜,其中所述纳米孔源自噬菌体dna包装马达的连接子蛋白。
在一些实施方式中,共聚物膜是三嵌段或二嵌段共聚物膜。由于纳米孔插入共聚物膜的方式,在一些实施方案中,包含纳米孔的共聚物膜进一步包含一种或多种脂质。在一些实施方式中,共聚物膜是两亲性aba型三嵌段共聚物膜。例如,可以通过使极性介质与包含aba分子的非极性介质接触来形成膜,这导致在非极性-极性界面处围绕极性介质自发形成aba分子层。当两个这样的极性介质体积然后通过非极性介质被放在一起时,在第一和第二极性体积之间的界面处形成aba分子的稳定膜。合成的所得膜已显示是坚固的、稳定的并且比常规脂质系统更不易于受到来自洗涤剂和蛋白质的降解。该膜还能够承受施加在其上的较大的电势差。
本文还公开了包含源自噬菌体dna包装马达的连接子蛋白的纳米孔的共聚物膜阵列。在一个实施方案中,所述阵列适于插入适于检测通过阵列中的纳米孔的分析物的移位的装置中。可以提供纳米孔阵列以增加通量,并因此增加多核苷酸链的测量,例如在国际申请wo2014/064443中公开的,该申请通过引用整体并入本文。
还公开了一种装置,其包括共聚物膜阵列,其中插入了源自噬菌体dna包装马达的连接子蛋白的纳米孔,以及用于跨所述膜施加电压电势的工具和用于检测跨所述膜电变化的工具。在一个实施方案中,所述装置还包括流体系统,其被配置成可控地向所述膜供应待表征的样品。
还公开了一种表征目标分析物的方法,该方法包括使包含源自噬菌体dna包装马达的连接子蛋白的纳米孔的共聚物膜与目标分析物接触,使得目标分析物相对于纳米孔移动,并且当目标分析物相对于孔移动时进行一次或多次测量,从而测定分析物的存在、不存在或一种或多种特征。在一些实施例中,测量是电测量和/或光学测量。在一些实施方案中,目标分析物与医学病症相关。
在本文所述的各个方面的一些实施方案中,所述方法还可以包括,在施加跨膜电势时,检测响应于分析物(例如,通过聚合酶加工多核苷酸的副产物)穿过纳米孔的信号。在一些实施方案中,电势差可以由提供离子流的渗透不平衡驱动。在一些实施方案中,可以跨越位于纳米孔任一侧上的两个电极之间的纳米孔施加电势差。信号可以是电测量和/或光学测量。可能的电测量包括:电流测量、阻抗测量、隧穿或电子隧穿测量(ivanovap等,nanolett.2011jan12;11(1):279-85)和fet测量(国际申请wo2005/124888),例如电压fet测量。光学测量可以与电测量相结合(sonigv等,revsciinstrum.2010jan;81(1):014301)。测量可以是跨膜电流测量,诸如流经纳米孔的离子电流的测量。
或者,测量可以是指示通过通道的离子流的荧光测量,例如由heron等,j.am.chem.soc.,2009,131(5),1652-1653公开,或使用fet测量跨膜的电压。在一些实施方案中,所述方法可以进一步包括,在施加跨膜的电势时,随着多核苷酸被处理,检测通过纳米孔的离子电流。在一些实施方案中,所述方法可以使用膜片钳或电压钳进行。在一些实施方案中,所述方法可以使用电压钳来进行。电测量可以使用标准单通道记录器材进行,如stoddartd等,procnatlacadsci,12;106(19):7702-7,liebermankr等,jamchemsoc.2010;132(50):17961-72,和国际申请wo2000/28312中所述。或者,可以使用多通道系统进行电测量,例如国际申请wo2009/077734和国际申请wo2011/067559中所述,其全部内容通过引用并入本文。
本文还公开了表征目标分析物的方法,其涉及跨本文公开的纳米孔传感器施加电势,使纳米孔传感器与目标分析物接触,使得目标分析物通过纳米孔,和检测目标分析物对通过纳米孔传感器的电导率的影响。
在下面的附图和描述中阐述了本发明的一个或多个实施例的细节。本发明的其它特征、目的和优点将从说明书和附图以及权利要求书中显而易见。
附图说明
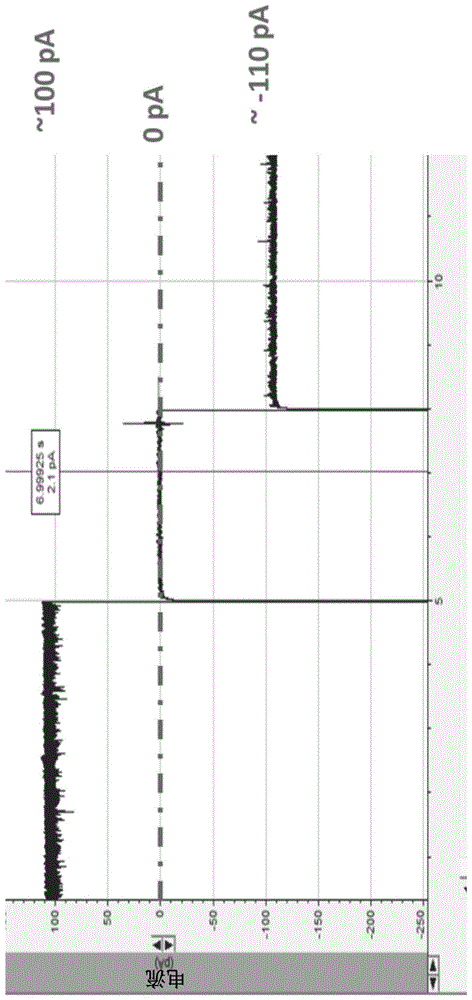
图1显示了使用minion电导缓冲液插入到脂质膜中的c-hisgp10/脂质体的单一通道。施加电压:+50mv或-50mv。
图2a到2d显示了在minion的通道256和267上的单一phi29通道插入。连接子插入之前通道256(图2a)和267(图2b)的电流约为0pa。单一通道插入是在不同电压下从通道256(图2c,施加的电压;100mv)和267(图2d,施加的电压:180mv)进行的。
图3a和3b显示了单一phi29连接子的三步门控以确认通道插入。图3a显示了在连接子插入之前minion的通道86的控制。图3b显示了对于通道86由100mv引起的单一phi29连接子的三步门控。
图4a和4b显示了肽通过minion的通道304的phi29连接子的单一通道的移位。图4a显示了在通道插入之前对通道304的控制。图4b显示肽通过位于通道304的单一phi29连接子移位。
图5a至5c显示了将噬菌体phi29dna包装马达的通道插入minion流动池的共聚物膜中。图5a显示了在50mv电压下phi29的单一通道的电流轨迹。图5b显示了通过电生理学测定用minion流动池缓冲液在±50mv的电压下测量的插入脂质双层膜的单一phi29通道。图5c示出了在100mv电压下单一phi29通道的三步门控。线表示施加的电压。
图6a和6b显示了tat肽在75mv(图6a)和100mv(图6b)下通过插入minion流动池的phi29通道的移位。线表示施加的电压。
图7a到7d示出了通过插入minion流动池的phi29通道在100mv下对tat肽(图7a)、r14肽(图7b)、mar肽(图7c)和p27肽(图7d)的不同肽的区分。线表示施加的电压。
具体实施方式
在更详细地描述本公开之前,应当理解,本公开不限于所描述的特定实施例,并且因此当然可以变化。还应理解,本文所用的术语仅用于描述特定实施方案的目的,而不旨在限制,因为本公开的范围将仅由所附权利要求书限制。
在提供数值范围的情况下,应理解为,除非上下文另有明确规定,否则每个中间数值均应达到该下限单位的十分之一;所述每个中间数值应为介于该范围的上限和下限之间以及在所述范围内的任何其它所述数值或中间数值。所述每个中间数值包括在本公开中。这些较小范围的上限和下限可以独立地包含在较小范围中,并且还应包括在本公开内容之内,但要遵守所述范围内任何明确排除的极限。在所述范围包括一个或两个极限的情况下,排除一个或两个那些所包括的极限的范围也包括在本公开中。
除非另外定义,否则本文所用的所有技术和科学术语具有与本公开所属领域的普通技术人员通常理解的相同的含义。尽管与本文所述的那些类似或等同的任何方法和材料也可用于本公开的实践或测试,但现在描述优选的方法和材料。
本说明书中引用的所有出版物和专利均通过引用并入本文,就像每个单独的出版物或专利具体地和单独地指明通过引用并入本文,并且通过引用并入本文以公开和描述与所引用的出版物相关的方法和/或材料。任何出版物的引用是因为其公开早于申请日,并且不应被解释为承认本公开由于在先公开而无权先于这样的出版物。此外,所提供的出版日期可能与实际出版日期不同,实际出版日期可能需要独立确认。
如本领域技术人员在阅读本公开后将显而易见的,本文描述和示出的每个单独实施例具有离散的部件和特征,其可容易地与其它若干实施例中的任一个的特征分离或组合,而不脱离本公开的范围或精神。任何列举的方法可以以列举的事件的顺序或以逻辑上可能的任何其它顺序来执行。
除非另有说明,本公开的实施方案将采用化学、生物学等技术,这些技术在本领域的技术范围内。
提出以下实施例以便为本领域普通技术人员提供如何进行本文公开和要求保护的方法和使用本文公开和要求保护的探针的完整公开和描述。已经努力确保关于数字(例如,数量、温度等)的准确性,但是应当考虑一些误差和偏差。除非另有说明,份数是重量份数,温度是℃并且压力是大气压或接近大气压。标准温度和压力定义为20℃和1个大气压。
在详细描述本公开的实施方案之前,应当理解,除非另有说明,本公开不限于特定材料、试剂、反应材料、制造工艺等,因为它们可以变化。还应理解,本文所用的术语仅是为了描述具体实施方案的目的,而不是为了限制。在本公开中,在逻辑上可能的情况下,还可以以不同的顺序执行步骤。
必须注意,如在说明书和所附权利要求中所使用的,单数形式“一”、“一个”和“该”包括复数指代物,除非上下文另外清楚地指明。
纳米孔
本文公开了纳米孔,其包含噬菌体dna包装马达的连接子蛋白通道。这些纳米孔可以并入本文公开的共聚物膜中。噬菌体dna包装马达的连接子蛋白通道不是膜蛋白。在本领域中已经描述了将马达通道插入脂质体和脂质双层膜中。但是将它们插入共聚物膜中在此首次描述。噬菌体dna包装马达的连接子蛋白通道首先插入脂质体中。脂质体可以与脂质膜融合,如前所述。本文公开了包含连接子蛋白通道的脂质体与共聚物膜而不是脂质膜的融合体。施加的跨共聚物膜的电势导致脂质体与共聚物膜融合。如本文所公开的,脂质体中的连接子蛋白通道可以插入共聚物膜中,使得它们形成跨膜通道。如本文所公开的,这些通道允许分析物进入并跨越共聚物膜。
纳米孔可以以与用于分析分析物与单独纳米孔的相互作用的装置兼容的格式插入到膜中,例如其中各自包含单个纳米孔的多个膜存在于阵列中的格式,其中装置被设置成单独地检测流过每个纳米孔的电流的变化。纳米孔可以用作传感器,用于表征目标分析物,例如多核苷酸或多肽。本文还公开了由噬菌体dna包装马达的连接子蛋白通道制备纳米孔传感器的方法。纳米孔的这种使用允许高通量、同时检测许多类型的癌症生物标记物,并且使得能够进行诸如蛋白质测序的应用。
本文首次描述了将病毒dna包装马达的连接子蛋白通道插入共聚物膜中。该共聚物膜至少稳定几个月,并且比以前插入了病毒dna包装马达的连接子蛋白通道的脂质双层耐受更高的电压。在用于检测装置的共聚物膜中插入孔的标准方法中,将孔的亚基以纯化形式悬浮于含有共聚膜例如三嵌段共聚物膜的溶液中,使得孔扩散到膜中,并通过与膜结合而插入并组装成功能状态。然而,为了将源自病毒dna包装马达的连接子蛋白通道的纳米孔插入到共聚物膜中,在施加的电势下,将插入纳米孔的脂质体与共聚物膜融合。
dna包装马达连接子蛋白
所述纳米孔源自病毒dna包装马达的连接子蛋白。连接子蛋白可以是噬菌体dna包装马达的连接子蛋白。连接子蛋白可以是野生型蛋白,或者与野生型连接子蛋白相比可以是修饰的。在一个实施方案中,修饰的蛋白是连接子蛋白的截短形式,例如连接子蛋白的片段或部分,和/或修饰的蛋白可以在连接子蛋白的一个或多个亚基的一个或两个末端包含额外的氨基酸,并且可以在连接子蛋白的氨基酸序列内包含一个或多个氨基酸取代、缺失或添加。
在一些实施方案中,纳米孔包含连接子蛋白的孔形成区。纳米孔可以例如包含连接子蛋白的一个或多个截短的亚基。在一个实施方案中,修饰孔形成区以改变纳米孔的通道的一种或多种性质。
在一些实施方案中,纳米孔包含完整连接子蛋白。例如,连接子蛋白仅包含连接子蛋白的全长亚基。
在一些实施方案中,纳米孔是由六个或更多个连接子蛋白亚基,例如7、8、9、10、11或12个亚基形成的多聚体蛋白。例如,纳米孔可以是十二聚体蛋白。一个或多个,例如2、3、4、5、6、7、8、9、10、11或12个亚基可以被修饰。在一个实施方案中,一个或多个亚基可以在c端和/或n端被修饰,例如以增加纳米孔的一端或两端的亲水性。例如,一个或多个亚基可通过在c端和/或n端添加柔性接头和/或肽标签来修饰。在一些实施方案中,纳米孔由相同的亚基组成。
可以使用任何合适的接头,例如包含3至12个氨基酸的接头,例如4或5至10个,优选6至8个氨基酸的接头。接头中的氨基酸可以选自赖氨酸、丝氨酸、精氨酸、脯氨酸、甘氨酸和/或丙氨酸。合适的接头的实例包括但不限于以下:gggs、pggs、pggg、rppppp、rpppp、vgg、rppg、pppp、rppg、ppppppppp、rppg、ggg、gggg、ggggg和gggggg。
任何合适的肽标签可以用于促进连接子蛋白的纯化。例如,在一个实施例中,标签可以是strep标签。在一个实施方案中,strep标签具有8至11个氨基酸的长度和/或strep标签氨基酸序列包含基序hpq。strep标签可以例如包含氨基酸序列wshpqsek、wshpqfek、nwshpqfek、pwshpqfek或ggshpqfeg,或由其组成。该序列可以通过添加、缺失或取代一个或多个氨基酸,如2、3、4或5个氨基酸而改变,只要保持核心“hpq”基序。变体序列通常为8至11个氨基酸nwshpqfek、pwshpqfek和ggshpqfeg。
噬菌体的atp驱动马达将病毒的线性dsdna包装到预先形成的前衣壳(procapsid)中。该马达的蛋白质中心是截短锥结构,称为连接子,其允许dsdna在成熟期间进入并在感染期间离开。连接子具有由十二个gp10蛋白亚基形成的中心通道。纯化了噬菌体phi29的示例性未修饰的病毒dna包装马达连接子蛋白,并结晶学表征了其三维结构(例如guasch等,1998febslett.430:283;marais等,2008structure16:1267)。phi29通道具有3.6nm窄和6nm宽的末端,其大于大多数膜蛋白通道。因此,本文所述的许多优选实施方案涉及phi29dna包装马达连接子蛋白(例如基因库登记号ace96033)和/或其多肽亚基,包括其能够形成通道的片段、变体和衍生物(例如登记号gi29565762、gi31072023、gi66395194、gi29565739、gi157738604)。
虽然病毒的连接子蛋白具有很少的序列同源性并且分子量不同,但是存在显著的潜在结构相似性。特别地,尽管与phi29连接子具有很少序列同源性并且分子量不同,其它dsdna病毒(例如,t4、lambda、p22、p2、t3、t5和t7)的dna包装马达连接子蛋白表现出显著的潜在结构相似性(例如,bazinet等,1985annrev.microbiol.39:109-29)。
在某些实施方案中,考虑使用来自其它dsdna病毒的分离的病毒dna包装马达连接子蛋白,包括但不限于来自任何噬菌体lambda、p2、p3、p22、t3、t4、t5、spp1、h97和t7的分离的病毒dna包装马达连接子蛋白,例如分离的dsdna病毒dna包装马达连接子蛋白(例如t4(登记号np-049782)(driedonks等,1981jmolbiol152:641),lambda(登记号gi549295,gi6723246,gi15837315,gi16764273)(kochan等,1984jmolbiol174:433),spp1(登记号p54309),p22(登记号aaa72961)(cingolani等,2002jstructbiol139:46),p2(登记号np-046757),p3(nutter等,1972j.viral.10(3):560-2),t3(登记号caa35152)(carazo等,1986jl.ultrastructmolstructres94:105),t5(登记号aax12078,yp-006980;aas77191;aau05287),t7(登记号np-041995)(cerritelli等,1996jmolbiol285:299;agirrezabala等,2005jmolbiol347:895))。
不希望受理论的束缚,在这方面,据信与本文示例的phi29dna包装马达连接子蛋白类似,这些和其它dsdna病毒包装马达连接子蛋白,其已经基本上在结构上被表征,可以并入膜层中以形成孔,当以与phi29dna包装马达的连接子蛋白相同的方式跨膜施加电势时,通过该孔可以发生电导。因此,对于某些实施方案,本文关于phi29连接子蛋白的公开内容旨在说明考虑了使用任何此类其它分离的dsdna病毒dna包装马达连接子蛋白的相关实施方案。
phi29dna包装马达的连接子蛋白或来自另一种噬菌体dna包装马达的连接子蛋白可以根据本文的教导进行修饰。
如本文更详细描述的,分离的dna包装马达连接子蛋白多肽,包括已经被人工工程化以具有膜结合特性(例如,膜层中的稳定跨膜整合)和功能导电跨膜孔形成特性的此类多肽,可以用作癌症生物标记物的导电生物传感器。
如本文所述,phi29和其它分离的dsdna病毒dna包装马达蛋白连接子,包括其工程化和突变形式,例如融合蛋白,其保留其孔结构域并包含亲和结构域,可有用地并入膜层中以形成孔,从而当跨膜施加电势时,允许其用作导电通道。
修饰的分离的双链dna病毒dna包装马达蛋白连接子如phi29连接子可以被工程化以具有用于本公开的实施方案的期望结构,其中蛋白晶体学结构数据是容易获得的。已经开发了大规模生产和纯化phi29连接子的程序(guo等,2005;ibanez等,nucleicacidsres.12,2351-2365(1984),robinson等,nucleicacidsres.34,2698-2709(2006),xiao等,acsnano3,100-107(2009))。
公开了用于方法中并且包括在组合物中的是源自病毒dna包装马达连接子蛋白的多肽和融合蛋白,所述多肽和融合蛋白具有与本领域已知序列相同或相似的氨基酸序列区,或其片段或部分。例如,通过举例说明而非限制,考虑了突变噬菌体phi29病毒dna包装马达连接子蛋白[例如,基因库登记号ace96033]或工程化源自噬菌体phi29病毒dna包装马达连接子蛋白的多肽融合蛋白用于本文公开的组合物和方法,与本文公开的多肽以及此类多肽的部分具有至少80%、90%或95%相似性的多肽,其中突变体或工程化源自phi29病毒dna包装马达连接子蛋白的多肽的这些部分通常包含至少150、175、200、225、250、275,包括至少240、260、280、285、290、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330个或更多氨基酸。
以类似的方式,某些其它实施方案考虑了其它突变双链dna噬菌体病毒马达连接子蛋白,例如突变形式的噬菌体t4dna包装马达连接子蛋白多肽,lambda噬菌体dna包装马达连接子蛋白多肽(登记号gi549295、gi6723246、gi15837315、gi16764273),噬菌体spp1dna包装马达连接子蛋白多肽(登记号p54309),噬菌体p22dna包装马达连接子蛋白多肽(登记号aaa72961),噬菌体p2dna包装马达连接子蛋白多肽(登记号np-046757),噬菌体p3dna包装马达连接子蛋白多肽(nutter等,1972j.virol.10(3):560-2),噬菌体t3dna包装马达连接子蛋白多肽(登记号caa35152),噬菌体t5dna包装马达连接子蛋白多肽(登记号aax12078,yp006980;aas77191;aau05287),噬菌体t7dna包装马达连接子蛋白多肽(登记号np041995)和噬菌体hk97dna包装马达连接子蛋白多肽(登记号np_037699)。例如,考虑了将任何这些噬菌体病毒dna包装马达连接子蛋白的突变体或任何这些噬菌体病毒dna包装马达连接子蛋白的工程化形式,其可以是多肽融合蛋白,用于所公开的组合物和方法中,以及与本文公开的多肽和此类多肽的片段具有至少80%、90%或95%相似性的多肽。突变或工程化的源自病毒dna包装马达连接子蛋白的多肽的“片段”通常包含至少150、175、200、225、250、275,包括至少240、260、280、285、290、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330个或更多氨基酸。
术语“片段”、“衍生物”和“类似物”在涉及病毒dna包装马达连接子蛋白或多肽时,是指本文所述的突变源自病毒dna包装马达连接子蛋白的多肽的变体,或包含此类多肽的融合蛋白,其基本上保留与此类多肽相同的生物功能或活性。因此,类似物包括前蛋白,其可通过切割前蛋白部分而激活,以产生活性病毒dna包装马达连接子多肽,在优选的实施方案中,其可并入膜层中以形成孔,当跨膜施加电势时,通过该孔可发生电导,和/或可能够自组装成同十二聚体病毒dna包装马达连接子蛋白,例如可形成这样的孔以获得含导电通道的膜。
本文所述的源自病毒dna包装马达连接子蛋白的多肽的片段、衍生物或类似物,包括多肽或融合蛋白或其结构域或片段,可以是(i)其中一个或多个氨基酸残基被保守或非保守氨基酸残基(优选保守氨基酸残基)取代,并且这种取代的氨基酸残基可以是或可以不是由遗传密码编码的所述片段、衍生物或类似物,或(ii)其中一个或多个氨基酸残基包括取代基的所述片段、衍生物或类似物,或(iii)其中额外的氨基酸融合到突变的源自病毒dna包装马达连接子蛋白的多肽上的所述片段、衍生物或类似物,包括用于检测或特异性功能改变突变的或工程化的源自病毒dna包装马达连接子蛋白的多肽或前蛋白序列的所述片段、衍生物或类似物。根据本文的教导,认为这些片段、衍生物和类似物在本领域技术人员的范围内。
如本领域已知的,通过将多肽的氨基酸序列和其保守氨基酸取代与第二多肽的序列进行比较来确定两个多肽之间的“相似性”。根据本发明公开的实施方式,编码多肽的核酸片段可用于合成编码突变或工程化的源自病毒dna包装马达连接子蛋白的多肽的全长核酸。如本文所用,“%同一性”是指当两个或多个多肽进行比对并使用空位(gapped)blast算法(例如,altschul等,1997nucl.ac.res.25:3389)分析其序列时,位于相应氨基酸残基位置的相同氨基酸的百分比,所述空位blast算法根据国家卫生研究院/ncbi数据库(国家生物技术信息中心,bethesda,md)提供的默认权重对序列空位和序列错配进行加权。
术语“分离的”是指从材料的原始环境(例如,如果该材料是天然存在的,则是天然环境)中移出该材料。例如,存在于活的动物中的或完整的天然存在的病毒中的天然存在的核酸或多肽不是分离的,但是从天然系统中的一些或全部共存材料分离的相同核酸或多肽是分离的。这样的核酸可以是载体的一部分和/或这样的核酸或多肽可以是组合物的一部分,并且仍然是分离的,因为这样的载体或组合物不是其天然环境的一部分。分离噬菌体马达蛋白的连接子蛋白的方法是本领域已知的。用于本文所述方法和组合物的噬菌体马达蛋白的连接子蛋白可以使用本领域熟知的方法重组产生。
术语“基因”是指参与产生多肽链的dna片段;它包括编码区“前导区和尾随区”之前和之后的区域以及各个编码片段(外显子)之间的间插序列(内含子)。
聚合物膜
根据本发明可以使用任何聚合物膜。合适的聚合物膜是本领域公知的。膜优选地是两亲性层。两亲性层是由两亲性分子如磷脂形成的层,所述两亲性分子具有至少一个亲水部分和至少一个亲脂或疏水部分。嵌段共聚物是聚合材料,其中两种或更多种单体亚基聚合在一起以产生单一聚合物链。嵌段共聚物通常具有由每个单体亚基贡献的性质。然而,嵌段共聚物可具有由单独的亚基形成的聚合物不具有的独特性质。嵌段共聚物可以被工程化成使得单体亚基之一是疏水性的(即亲脂性的),而其它亚基在水性介质中是亲水性的。在这种情况下,嵌段共聚物可具有两亲性性质,并且可形成模拟生物膜的结构。嵌段共聚物可以是二嵌段(由两个单体亚基组成),但也可以由多于两个单体亚基构成,以形成表现为两亲物的更复杂的排列。共聚物可以是三嵌段、四嵌段或五嵌段共聚物。
在一些实施方式中,共聚物膜是两亲性aba型三嵌段共聚物膜。aba分子的特别稳定的膜在wo2014064444a1中有所描述,其全部内容通过引用并入本文,以教导这些膜及其在形成纳米孔序列中的用途。
两亲性分子可以是合成的或天然存在的。非天然存在的两亲物和形成单层的两亲物是本领域已知的,包括例如嵌段共聚物(gonzalez-perez等,langmuir,2009,25,10447-10450)。嵌段共聚物是聚合材料,其中两种或更多种单体亚基聚合在一起以产生单一聚合物链。嵌段共聚物通常具有由每个单体亚基贡献的性质。然而,嵌段共聚物可具有由单独的亚基形成的聚合物不具有的独特性质。嵌段共聚物可以被工程化成使得单体亚基之一是疏水性的(即亲脂性的),而其它亚基在水性介质中是亲水性的。在这种情况下,嵌段共聚物可具有两亲性性质,并且可形成模拟生物膜的结构。嵌段共聚物可以是二嵌段(由两个单体亚基组成),但也可以由多于两个单体亚基构成,以形成表现为两亲物的更复杂的排列。共聚物可以是三嵌段、四嵌段或五嵌段共聚物。膜优选为三嵌段共聚物膜。
古生细菌双极四醚脂质是天然存在的脂质,其被构造为使得脂质形成单层膜。这些脂质通常在极端微生物中发现,所述极端微生物在恶劣的生物环境中存活,嗜热菌,嗜盐菌和嗜酸菌。据信它们的稳定性来自最终双层的融合性质。通过产生具有一般基序亲水-疏水-亲水的三嵌段聚合物来构建模拟这些生物实体的嵌段共聚物材料是直接的。该材料可形成单体膜,其行为类似于脂质双层,并涵盖从囊泡到层状膜的一系列相行为。由这些三嵌段共聚物形成的膜具有优于生物脂质膜的几个优点。因为合成了三嵌段共聚物,所以可以仔细地控制精确的结构以提供形成膜和与孔和其它蛋白质相互作用所需的正确的链长和性质。
嵌段共聚物还可由未归类为脂质子材料的亚基构造;例如,疏水性聚合物可以由硅氧烷或其它非烃基单体制成。嵌段共聚物的亲水性亚区段也可以具有低蛋白结合性质,这允许产生当暴露于原始生物样品时高度耐受的膜。该头基单元也可以源自非经典脂质头基。
三嵌段共聚物膜与生物脂质膜相比还具有增加的机械和环境稳定性,例如高得多的操作温度或ph范围。嵌段共聚物的合成性质提供了定制用于广泛应用的基于聚合物膜的平台。
该膜最优选地是在国际申请号wo2014/064443或wo2014/064444中公开的膜中的一种,这些申请通过引用整体结合到本文中。
两亲性分子可以是化学修饰的或功能化的,以促进多核苷酸的偶联。两亲性层可以是单层或双层。两亲性层通常是平面的。两亲性层可以是弯曲的。两亲性层可以是被支持的。
两亲性膜通常是天然可移动的,基本上充当具有约10-8cms-1的脂质扩散速率的二维流体。这意味着孔和偶联的多核苷酸通常可以在两亲性膜内移动。
在一些实施方案中,共聚物膜可包含一种或多种添加剂,例如以影响层的性质。合适的添加剂包括但不限于脂肪酸,例如棕榈酸、肉豆蔻酸和油酸;脂肪醇,例如棕榈醇、肉豆蔻醇和油醇;甾醇,如胆固醇、麦角固醇、羊毛甾醇、谷甾醇和豆甾醇;溶血磷脂,如1-酰基-2-羟基-sn-甘油基-3-磷酸胆碱;和神经酰胺。共聚物膜可以例如包含脂质,所述脂质是与共聚物膜融合以将纳米孔插入膜中的脂质体的组分。
共聚物膜可以包含在固态层中。固态层可以由有机和无机材料形成,其包括但不限于微电子材料、绝缘材料例如si3n4、al2o3和sio、有机和无机聚合物例如聚酰胺、塑料例如特氟隆
纳米孔并入聚合物膜中
可以通过首先将如本文提供的分离的病毒dna包装马达连接子蛋白并入到脂质体中来形成当前描述的包含导电通道的膜,然后所述脂质体可以通过人工膜融合操作将膜整合的连接子提供给聚合物膜系统。
如本文所用,术语“脂质体”是指具有至少一个脂质双层的脂质囊泡。脂质体最通常由磷脂,尤其是磷脂酰胆碱组成,但也可包括其它脂质。脂质体的实例包括多层囊泡(mlv,具有几个层状相脂质双层)、小单层脂质体囊泡(suv,具有一个脂质双层)、大单层囊泡(luv)和脂质卷(cochleate)囊泡。脂质体的另一种形式是多囊脂质体,其中一个囊泡含有一个或多个较小的囊泡。脂质体可以通过破坏生物膜(例如通过超声处理)来制备,脂质体也可以通过超声处理两亲性脂质(例如磷脂)在水中的分散体来产生。另一种选择是mozafari法,该方法包括使用加热形成脂质体,同时搅拌以使各成分均匀分布。
在公开的方法中,纳米孔首先插入脂质体中。脂质体可以例如是稳定的、巨大的和/或单层脂质体。
脂质体可以例如通过超声处理、挤出或mozafari法(colas等(2007)micron38:841-847)形成。
脂质体可以包含脂质双层。脂质双层由两个相对的脂质层形成。排列两层脂质,使得它们的疏水性尾基彼此面对,形成疏水性内部。脂质的亲水性头基在双层的每一侧上面向外面朝向水性环境。双层可以存在于许多脂质相中,包括但不限于液体无序相(流体层状)、液体有序相、固体有序相(层状凝胶相、交叉凝胶相(interdigitatedgelphase))和平面双层晶体(层状亚凝胶相、层状结晶相)。
可以使用形成脂质双层的任何脂质组合物。选择脂质组合物,使得形成具有所需性质的脂质双层,所述所需性质例如表面电荷、支持膜蛋白的能力、填充密度(packingdensity)或机械性质。脂质组合物可包含一种或多种不同的脂质。例如,脂质组合物可以含有至多100种脂质。脂质组合物优选含有1至10种脂质。脂质组合物可包含天然存在的脂质和/或人工脂质。
脂质通常包含头基、界面部分和两个疏水性尾基,所述两个疏水性尾基可以相同或不同。合适的头基包括但不限于中性头基,例如二酰基甘油酯(dg)和神经酰胺(cm);两性离子头基,如磷脂酰胆碱(pc)、磷脂酰乙醇胺(pe)和鞘磷脂(sm);带负电荷的头基,例如磷脂酰甘油(pg)、磷脂酰丝氨酸(ps)、磷脂酰肌醇(pi)、磷酸(pa)和心磷脂(ca);和带正电荷的头基,例如三甲基铵-丙烷(tap)。合适的界面部分包括但不限于天然存在的界面部分,例如基于甘油或基于神经酰胺的部分。合适的疏水性尾基包括但不限于饱和烃链,例如月桂酸(正-十二烷酸)、肉豆蔻酸(正-十四烷酸)、棕榈酸(正-十六烷酸)、硬脂酸(正-十八烷酸)和花生酸(正-二十烷酸);不饱和烃链,如油酸(顺-9-十八烷酸);和分支烃链,例如植烷酰基。不饱和烃链中链的长度以及双键的位置和数目可以变化。分支烃链中链的长度和分支如甲基的位置和数目可以变化。疏水性尾基可以作为醚或酯连接到界面部分。脂质可以是分枝菌酸。
脂质也可以是化学修饰的。脂质的头基或尾基可以是化学修饰的。其头基已经被化学修饰的合适的脂质包括但不限于peg修饰的脂质,例如1,2-二酰基-sn-甘油基-3-磷酸乙醇胺-n-[甲氧基(聚乙二醇)-2000];功能化peg脂质,如1,2-二硬脂酰基-sn-甘油基-3磷酸乙醇胺-n-[生物素基(聚乙二醇)2000];和经修饰用于缀合的脂质,例如1,2-二油酰基-sn-甘油基-3-磷酸乙醇胺-n-(琥珀酰基)和1,2-二棕榈酰基-sn-甘油基-3-磷酸乙醇胺-n-(生物素基)。其尾基已经被化学修饰的合适的脂质包括但不限于可聚合的脂质,例如1,2-双(10,12-二十三碳二炔酰基)-sn-甘油基-3-磷酸胆碱(1,2-bis(10,12-tricosadiynoyl)-sn-glycero-3-phosphocholine);氟化脂质,如1-棕榈酰基-2-(16-氟棕榈酰基)-sn-甘油基-3-磷酸胆碱;氘化脂质,如1,2-二棕榈酰基-d62-sn-甘油基-3-磷酸胆碱;和醚连接的脂质,例如1,2-二-o-植烷基-sn-甘油基-3-磷酸胆碱。脂质可以是化学修饰的或功能化的以促进多核苷酸的偶联。
脂质组合物通常包含一种或多种影响层性质的添加剂。合适的添加剂包括但不限于脂肪酸,例如棕榈酸、肉豆蔻酸和油酸;脂肪醇,例如棕榈醇、肉豆蔻醇和油醇;甾醇,如胆固醇、麦角固醇、羊毛甾醇、谷甾醇和豆甾醇;溶血磷脂,如1-酰基-2-羟基-sn-甘油基-3-磷酸胆碱;和神经酰胺。
在一些实施方案中,所述方法包括将噬菌体dna包装马达的连接子蛋白通道并入脂质体中,使共聚物膜与脂质体接触,并通过跨共聚物膜施加电压诱导脂质体-聚合物融合。在一个实施方案中,施加的电位为50mv至500mv,100mv至360mv,200mv至400mv,300mv至350mv,或300mv至325mv。可以使用任何合适的方法,使用电压控制来控制膜中纳米孔的插入。电压辅助的孔插入是已知的,并且描述于us9,797013和wo2018/096348中,在此通过引用将其全部内容并入本文。minknow是由oxfordnanoporetechnologiesltd提供的软件包,其在本文所述的方法中使用和引用以施加跨膜的电势差并促进孔插入。在电压辅助的孔插入的方法中,可施加初始电压,然后逐渐增加或随时间逐步增加到最终电压。在一个实施例中,施加初始电压(例如,从50到100mv),并且电压每10秒增加5mv,直到500mv的电压。在另一个实施例中,施加初始电压(例如,从50到100mv),并且电压每10秒增加5mv,直到360mv的电压。
脂质体可以用干燥的dphpc(1,2-二植烷酰基-sn-甘油基-3-磷酸胆碱)制备。纯化的连接子蛋白和再水化缓冲液可以加入到制备的脂质体中。所得溶液含有包含连接子蛋白的脂质体。然后,这些脂质体可用于通过脂质体融合将连接子蛋白插入到共聚物膜中。
表征分析物的方法
还公开了使用所公开的纳米孔表征目标分析物的方法。所述方法包括使所述目标分析物与公开的纳米孔系统接触,使得所述目标分析物移动通过所述纳米孔。然后使用本领域已知的标准方法,当分析物相对于纳米孔移动时,测量目标分析物的一个或多个特征。优选地,当分析物移动通过纳米孔时测量目标分析物的一个或多个特征。这些步骤可以在施加跨过纳米孔的电势的情况下进行。例如,所施加的电势可以是电压电势、化学电势或电势。化学电势的一个实例是跨两亲性层的盐梯度。
可以进行各种不同类型的测量。这包括但不限于:电测量和光学测量。可能的电测量包括:电流测量、阻抗测量、隧穿测量(ivanovap等,nanolett.2011jan12;11(1):279-85)和fet测量(wo2005/124888)。光学测量可以与电测量相结合(sonigv等,revsciinstrum.2010jan;81(1):014301)。测量可以是跨膜电流测量,例如测量流经孔的离子电流。在一些情况下,利用场效应晶体管(fet)纳米孔传感器进行测量。在一些情况下,利用集成隧穿电极(integratedtunnelingelectrode)进行测量。
在一些实施方案中,所述方法涉及当分析物相对于纳米孔移动时测量通过纳米孔的电流,其中所述电流指示目标分析物的一种或多种特征,并由此表征目标分析物。在一些实施方案中,目标分析物是目标多核苷酸,并且所述方法涉及使目标多核苷酸与共聚物膜中存在的公开的纳米孔和多核苷酸结合蛋白接触,使得所述蛋白控制目标多核苷酸通过纳米孔的运动,然后测量当多核苷酸相对于纳米孔运动时通过纳米孔的电流,其中所述电流指示目标多核苷酸的一种或多种特征,从而表征目标多核苷酸。
所公开的方法可用于表征目标分析物。在一些实施方案中,目标分析物是金属离子、无机盐、聚合物、氨基酸、肽、多肽、蛋白质、核苷酸、寡核苷酸、多核苷酸、染料、漂白剂、药物、诊断试剂、消遣性毒品(recreationaldrug)、爆炸物或环境污染物。在一些实施方案中,分析物是氨基酸、肽、多肽和/或蛋白质。氨基酸、肽、多肽或蛋白质可以是天然存在的或非天然存在的。多肽或蛋白质可包括合成的或修饰的氨基酸在其中。许多不同类型的氨基酸修饰是本领域已知的。蛋白质可以是酶、抗体、激素、生长因子或生长调节蛋白,例如细胞因子。细胞因子可以选自白介素,如ifn-1、il-1、il-2、il-4、il-5、il-6、il-10、il-12和il-13,干扰素,如il-γ,和其它细胞因子,如tnf-α。蛋白质可以是细菌蛋白质、真菌蛋白质、病毒蛋白质或源自寄生虫的蛋白质。
目标分析物可以是核苷酸、寡核苷酸或多核苷酸。所公开的方法可以包括测量选自以下的一个或多个特征:(i)目标多核苷酸的长度,(ii)目标多核苷酸的同一性,(iii)目标多核苷酸的序列,(iv)目标多核苷酸的二级结构和(v)目标多核苷酸是否被修饰。
核苷酸通常含有核碱基、糖和至少一个磷酸基。核碱基通常是杂环的。核碱基包括但不限于嘌呤和嘧啶,更具体地说是腺嘌呤、鸟嘌呤、胸腺嘧啶、尿嘧啶和胞嘧啶。糖通常是戊糖。核苷酸糖包括但不限于核糖和脱氧核糖。核苷酸通常是核糖核苷酸或脱氧核糖核苷酸。核苷酸通常含有单磷酸、二磷酸或三磷酸。磷酸可以连接在核苷酸的5'或3'侧。核苷酸包括但不限于腺苷一磷酸(amp)、腺苷二磷酸(adp)、腺苷三磷酸(atp)、鸟苷一磷酸(gmp)、鸟苷二磷酸(gdp)、鸟苷三磷酸(gtp)、胸苷一磷酸(tmp)、胸苷二磷酸(tdp)、胸苷三磷酸(ttp)、尿苷一磷酸(ump)、尿苷二磷酸(udp)、尿苷三磷酸(utp)、胞苷一磷酸(cmp)、胞苷二磷酸(cdp)、胞苷三磷酸(ctp)、5-甲基胞苷一磷酸、5-甲基胞苷二磷酸、5-甲基胞苷三磷酸、5-羟甲基胞苷一磷酸、5-羟甲基胞苷二磷酸、5-羟甲基胞苷三磷酸、环腺苷一磷酸(camp)、环鸟苷一磷酸(cgmp)、脱氧腺苷一磷酸(damp)、脱氧腺苷二磷酸(dadp)、脱氧腺苷三磷酸(datp)、脱氧鸟苷一磷酸(dgmp)、脱氧鸟苷二磷酸(dgdp)、脱氧鸟苷三磷酸(dgtp)、脱氧胸苷一磷酸(dtmp)、脱氧胸苷二磷酸(dtdp)、脱氧胸苷三磷酸(dttp)、脱氧尿苷一磷酸(dump)、脱氧尿苷二磷酸(dudp)、脱氧尿苷三磷酸(dutp)、脱氧胞苷一磷酸(dcmp)、脱氧胞苷二磷酸(dcdp)、脱氧胞苷三磷酸(dctp)、5-甲基-2'-脱氧胞苷一磷酸、5-甲基-2'-脱氧胞苷二磷酸、5-甲基-2'-脱氧胞苷三磷酸、5-羟甲基-2'-脱氧胞苷一磷酸、5-羟甲基-2'-脱氧胞苷二磷酸和5-羟甲基-2'-脱氧胞苷三磷酸。核苷酸优选选自amp、tmp、gmp、ump、damp、dtmp、dgmp或dcmp。核苷酸可以是脱碱基的(即缺少核碱基)。核苷酸可以含有额外的修饰。具体地,合适的修饰核苷酸包括但不限于2'氨基嘧啶(例如2'-氨基胞苷和2'-氨基尿苷)、2'-羟基嘌呤(例如、2'-氟嘧啶(例如2'-氟胞苷和2'氟尿苷)、羟基嘧啶(例如5'-α-β-硼烷尿苷)、2'-o-甲基核苷酸(例如2'-o-甲基腺苷、2'-o-甲基鸟苷、2'-o-甲基胞苷和2'-o-甲基尿苷)、4'-硫代嘧啶(例如4'-硫代尿苷和4'-硫代胞苷)和具有核碱基修饰的核苷酸(例如5-戊炔基-2'-脱氧尿苷、5-(3-氨基丙基)-尿苷和1,6-二氨基己基-n-5-氨基甲酰基甲基尿苷)。寡核苷酸是短核苷酸聚合物,其通常具有50个或更少的核苷酸,例如40个或更少、30个或更少、20个或更少、10个或更少或5个或更少。寡核苷酸可包含以上讨论的任何核苷酸,包括脱碱基和修饰的核苷酸。
多核苷酸可以是单链或双链的。在一些情况下,多核苷酸的至少一部分是双链的。单链多核苷酸可以具有与其杂交的一个或多个引物,因此包含双链多核苷酸的一个或多个短区域。引物可以是与目标多核苷酸相同类型的多核苷酸,或者可以是不同类型的多核苷酸。
多核苷酸可以是核酸,例如脱氧核糖核酸(dna)或核糖核酸(rna)。目标多核苷酸可以包含与一条dna链杂交的一条rna链。多核苷酸可以是本领域已知的任何合成核酸,例如肽核酸(pna)、甘油核酸(gna)、苏糖核酸(tna)、锁核酸(lna)或具有核苷酸侧链的其它合成聚合物。
可以使用该方法表征全部或仅部分的目标多核苷酸。目标多核苷酸可以是任何长度。例如,多核苷酸的长度可以是至少10、至少50、至少100、至少150、至少200、至少250、至少300、至少400或至少500个核苷酸对。多核苷酸的长度可以是1000或更多个核苷酸对、5000或更多个核苷酸对或100000或更多个核苷酸对。
所公开的方法通常在已知含有或怀疑含有目标分析物的样品上进行。或者,可以对样品进行该方法以确认一种或多种目标分析物的身份,所述目标分析物在样品中的存在是已知的或预期的。在一些实施方案中,样品是生物样品。该方法可以在体外(invitro)对从任何生物体或微生物获得或提取的样品进行。所述生物或微生物通常是古生的、原核的或真核的,并且通常属于五界之一:植物界、动物界、真菌界、原核生物界和原生生物界。该方法可以在体外对从任何病毒获得或提取的样品进行。在一些实施方案中,样品可以是流体样品,例如患者的体液。例如,样品可以是血液、血浆、血清、尿、淋巴液、唾液或羊水。通常,样品来源于人,但可选地,其可以来自另一种哺乳动物,例如来自商业养殖的动物,例如马、牛、羊或猪,或者可选地,可以是宠物,例如猫或狗。或者,植物来源的样品可以从商业作物获得,例如谷物、豆类、水果或蔬菜,例如小麦、大麦、燕麦、油菜、玉米、大豆、稻、香蕉、苹果、番茄、马铃薯、葡萄、烟草、豆类、小扁豆、甘蔗、可可、棉花。
样品也可以是环境样品。例如,样品可以是水样品,例如饮用水、海水或河水。
样品通常在测定之前进行处理,例如通过离心或通过穿过滤除不需要的分子或细胞的膜。样品可以在取样时立即测量。样品通常也可以在测定之前储存,例如低于-70℃。
在链测序中,dna在施加的电势下或逆着施加的电势移位通过纳米孔。对双链dna渐进或持续作用的外切核酸酶可用于纳米孔的顺侧(cisside)以在施加的电势下使剩余的单链通过,或用于纳米孔的反侧(transside)以在相反电势下使剩余的单链通过。同样,解旋双链dna的解旋酶也可以以类似的方式使用。也可使用聚合酶。对于需要针对所施加的电势的链移位的测序应用也存在可能性,但是dna必须首先在相反电势或无电势下被酶“捕获”。随着在结合之后电势转换回去,所述链将从顺侧通过纳米孔到反侧,并被电流保持在延伸的构象中。单链dna外切核酸酶或单链dna依赖性聚合酶可以作为分子马达,以受控的逐步方式,即反侧至顺侧,逆着施加的电势,将最近移位的单链拉回通过孔。
在一些实施方案中,表征目标多核苷酸的方法包括使目标序列与所公开的纳米孔和解旋酶接触。任何解旋酶都可以用于该方法中。解旋酶相对于孔可以以两种模式工作。首先,优选使用解旋酶进行该方法,使得其利用由施加的电压产生的场使目标序列移动通过孔。在这种模式下,dna的5'端首先被捕获在纳米孔中,然后酶将dna移入纳米孔,使得目标序列随电场穿过纳米孔,直到其最终移位到双层的反侧。或者,优选地进行该方法,使得解旋酶逆着由施加的电压产生的场使目标序列穿过纳米孔。在这种模式中,dna的3'末端首先被捕获在纳米孔中,并且酶移动dna通过该孔,使得目标序列逆着施加的电场被拉出纳米孔,直到最终被弹回到双层的顺侧。
在外切核酸酶测序中,外切核酸酶从目标多核苷酸的一端释放单个核苷酸,并且鉴定这些单个核苷酸。在另一个实施方案中,表征目标多核苷酸的方法包括使目标序列与公开的纳米孔和外切核酸酶接触。上述任何外切核酸酶都可用于本方法。如上文所讨论的,酶可以共价地附着到纳米孔。
外切核酸酶是通常锁在多核苷酸的一端并从该端一次一个核苷酸地消化序列的酶。外切核酸酶可以在5’至3’方向上或3’至5’方向上消化多核苷酸。外切核酸酶结合至多核苷酸的末端通常通过选择使用的酶和/或使用本领域已知的方法来确定。在多核苷酸的任一端的羟基或帽结构通常可以用于防止或促进外切核酸酶与多核苷酸的特定末端的结合。
该方法可包括使多核苷酸与外切核酸酶接触,使得从多核苷酸末端以允许如上所述表征或鉴定一部分核苷酸的速率消化核苷酸。用于这样做的方法在本领域中是公知的。例如,edman降解用于连续消化来自多肽末端的单个氨基酸,使得可以使用高效液相色谱(hplc)鉴定它们。在本方法中可以使用同源方法。
外切核酸酶发挥功能的速率通常比野生型外切核酸酶的最佳速率慢。在所公开的方法中,外切核酸酶活性的合适速率包括每秒0.5至1000个核苷酸、每秒0.6至500个核苷酸、每秒0.7至200个核苷酸、每秒0.8至100个核苷酸、每秒0.9至50个核苷酸或每秒1至20或10个核苷酸的消化。速率优选为1、10、100、500或1000个核苷酸每秒。外切核酸酶活性的合适速率可以以多种方式实现。例如,可以使用具有降低的最佳活性速率的变体外切核酸酶。
所公开的方法可以使用适合于研究膜/孔系统的任何设备来进行,在所述膜/孔系统中纳米孔被插入共聚物膜中。所述方法可以包括当分析物,例如目标多核苷酸,相对于纳米孔移动时,测量通过纳米孔的电流。因此,所述设备还可以包括能够施加电势并测量跨所述膜和纳米孔的电信号的电路。所述方法可以使用膜片钳或电压钳进行。
所公开的方法可以涉及当分析物诸如目标多核苷酸相对于纳米孔移动时测量通过纳米孔的电流。用于测量通过跨膜蛋白纳米孔的离子电流的合适条件是本领域已知的。所使用的电压通常为+2v至-2v,通常为-400mv至+400mv。所用电压可以在具有选自-400mv、-300mv、-200mv、-150mv、-100mv、-50mv、-20mv和0mv的下限和独立地选自+10mv、+20mv、+50mv、+100mv、+150mv、+200mv、+300mv和+400mv的上限的范围内。所用电压可以在100mv至240mv的范围内,例如120mv至220mv。通过使用增加的施加电势,可以增加纳米孔在不同核苷酸之间的辨别。
该方法通常在电荷载体存在下进行,所述电荷载体例如金属盐,例如碱金属盐,卤化物盐,例如氯化物盐,例如碱金属氯化物盐。电荷载体可以包括离子液体或有机盐,例如四甲基氯化铵、三甲基苯基氯化铵、苯基三甲基氯化铵或1-乙基-3-甲基咪唑鎓氯化物。在上述示例性设备中,盐存在于腔室中的水溶液中。通常使用氯化钾(kcl)、氯化钠(nacl)或氯化铯(cscl)。优选kcl。盐浓度可以是饱和的。盐浓度可以是3m或更低,通常为0.1至2.5m、0.3至1.9m、0.5至1.8m、0.7至1.7m、0.9至1.6m或1m至1.4m。盐浓度可以是150mm至1m。该方法可以使用至少0.3m,例如至少0.4m、至少0.5m、至少0.6m、至少0.8m、至少1.0m、至少1.5m、至少2.0m、至少2.5m或至少3.0m的盐浓度进行。高盐浓度提供高信噪比,并允许在正常电流波动的背景下鉴定指示核苷酸存在的电流。
所述方法可以在缓冲液的存在下进行。在上述示例性设备中,缓冲液存在于腔室中的水溶液中。任何合适的缓冲液都可以用于该方法中。通常,缓冲液是hepes。另一种合适的缓冲液是tris-hcl缓冲液。所述方法通常在4.0至12.0、4.5至10.0、5.0至9.0、5.5至8.8、6.0至8.7或7.0至8.8或7.5至8.5的ph下进行。
所述方法可以在0℃至100℃、15℃至95℃、16℃至90℃、17℃至85℃、18℃至80℃、19℃至70℃或20℃至60℃的温度下进行。所述方法可以在室温下进行。所述方法任选地在支持酶功能的温度,诸如约37℃下进行。
所述方法可以在游离核苷酸或游离核苷酸类似物和促进多核苷酸结合蛋白作用的酶辅因子,如解旋酶或外切核酸酶存在下进行。所述游离核苷酸可以是上述单个核苷酸中的任何一个或多个。游离核苷酸包括但不限于腺苷一磷酸(amp)、腺苷二磷酸(adp)、腺苷三磷酸(atp)、鸟苷一磷酸(gmp)、鸟苷二磷酸(gdp)、鸟苷三磷酸(gtp)、胸苷一磷酸(tmp)、胸苷二磷酸(tdp)、胸苷三磷酸(ttp)、尿苷一磷酸(ump)、尿苷二磷酸(udp)、尿苷三磷酸(utp)、胞苷一磷酸(cmp)、胞苷二磷酸(cdp)、胞苷三磷酸(ctp)、环腺苷一磷酸(camp)、环鸟苷一磷酸(cgmp)、脱氧腺苷一磷酸(damp)、脱氧腺苷二磷酸(dadp)、脱氧腺苷三磷酸(datp)、脱氧鸟苷一磷酸(dgmp)、脱氧鸟苷二磷酸(dgdp)、脱氧鸟苷三磷酸(dgtp)、脱氧胸苷一磷酸(dtmp)、脱氧胸苷二磷酸(dtdp)、脱氧胸苷三磷酸(dttp)、脱氧尿苷一磷酸(dump)、脱氧尿苷二磷酸(dudp)、脱氧尿苷三磷酸(dutp)、脱氧胞苷一磷酸(dcmp)、脱氧胞苷二磷酸(dcdp)、脱氧胞苷三磷酸(dctp)。酶辅因子是允许解旋酶发挥功能的因子。在一些实施方案中,酶辅因子是二价金属阳离子,例如mg2+、mn2+、ca2+或co2+。
本文还公开了用于表征例如测序目标多核苷酸的试剂盒。所述试剂盒包含本文公开的纳米孔系统和多核苷酸结合蛋白,例如解旋酶或外切核酸酶。所公开的试剂盒可以另外包含一种或多种其它试剂或仪器,其使得能够进行上述任何实施方案。这样的试剂或仪器包括以下的一种或多种:合适的缓冲液(水溶液)、从对象获得样品的工具(例如容器或包含针的仪器)、扩增和/或表达多核苷酸序列或电压或膜片钳设备的工具。试剂盒还可以任选地包含使试剂盒能够用于所公开的方法或关于该方法可以用于哪些患者的细节的说明书。试剂盒可以任选地包含核苷酸。
本文还公开了用于表征例如测序样品中的目标多核苷酸的设备。所述设备可以包括共聚物膜,所述共聚物膜含有多个本文公开的纳米孔和多个多核苷酸结合蛋白,例如解旋酶或外切核酸酶。该设备可以是用于分析物分析的任何常规设备,例如阵列或芯片。在一些实施方案中,所述设备可以是wo2010/122293、wo2011067559或wo2000/028312中描述的任何装置,这些文献通过引用整体结合到本文中。
具体地,公开了包含共聚物膜阵列的装置,所述共聚物膜包含源自噬菌体dna包装马达的连接子蛋白的纳米孔。该装置还可以包括用于跨膜施加电压电势的工具和用于检测跨膜电变化的工具。所述装置可以包含多个通道,其中每个通道包含共聚物膜,其中所述共聚物膜插入了源自噬菌体dna包装马达的连接子蛋白的纳米孔,和用于跨所述膜施加电压电势的工具和用于检测跨所述膜电变化的工具。该装置还可以包括流体系统,其被配置成可控地向膜供应待表征的样品。
以下是本文公开的发明的各个方面:
1.将噬菌体phi29、t3、t4、t5、t7、spp1和hk97dna包装马达的连接子蛋白通道插入来自oxfordnanoporetechnologies的minion。
2.通过两步法将噬菌体phi29、t3、t4、t5、t7、spp1和hk97dna包装马达的连接子蛋白通道插入minion聚合物膜中:第一步是将连接子蛋白插入脂质体,以及第二步是在所述minion中将脂质体与聚合物膜融合。
3.通过观察三步通道门控来确认phi29通道插入minion的方法。
4.使用含有噬菌体phi29、t3、t4、t5、t7、spp1和hk97dna包装马达的连接子蛋白通道的minion芯片进行高通量dna测序。
5.使用含有噬菌体phi29、t3、t4、t5、t7、spp1和hk97dna包装马达的连接子蛋白通道的minion芯片进行rna测序。
6.使用含有噬菌体phi29、t3、t4、t5、t7、spp1和hk97dna包装马达的连接子蛋白通道的minion芯片进行检测抗原、抗体和噬菌体展示肽以诊断癌症、病毒感染、细菌感染和其它疾病。
7.使用含有噬菌体phi29、t3、t4、t5、t7、spp1和hk97dna包装马达的连接子蛋白通道的minion芯片进行检测mirna以诊断癌症和其它疾病。
已经描述了本发明的多个实施例。然而,应当理解,在不背离本发明的精神和范围的情况下,可以进行各种修改。因此,其它实施例也在所附权利要求的范围内。
实施例
实施例1:将噬菌体phi29dna包装马达的连接子蛋白通道插入共聚物膜中。
图1-4显示了将噬菌体phi29dna包装马达的连接子蛋白通道插入到与minion装置一起使用的共聚物膜中。下列方法同样适用于本文所述的其它连接子蛋白通道,例如噬菌体p2、p3、p22、t3、t4、t5、spp1、h97、t7、g20c和lambda。
材料和方法:
phi29蛋白纯化方法:
phi29连接子的克隆、表达和纯化在以前的出版物(haque等,2013,nat.protoc.,8,373-392;wendell等,2009,nat.nanotechnol.,4,765-772)中有描述。简言之,将含有编码phi29连接子蛋白通道的单个亚基的基因10(gp10)的构建质粒转化到大肠杆菌hms174(de3)或bl21(de3)中。将含有phi29质粒的大肠杆菌在500mlluriabroth培养基中于37℃培养至od600达到0.5-0.6。在收获前用iptg(终浓度:0.5mm)诱导phi29蛋白表达额外3小时。将沉淀重悬于缓冲液(tris-hcl0.1m,nacl0.5m,atp10mm,甘油14.4%,咪唑5mm)中,裂解细胞。回收可溶性蛋白,通过离心收集,并通过0.45μm膜过滤器,然后上样到填充有his
phi29连接子并入脂质体中:
从avantipolarlipids获得1,2-二植烷酰基-sn-甘油基-3-磷酸胆碱(dphpc)。phi29蛋白脂质体(proteoliposome)的制备方法是先前方案(danilo,1997,crcpressllc;haque等,2013,nat.protoc.,8,373-392)的改进版本。简言之,使用旋转蒸发器蒸发掉含有1mgdphpc的氯仿,然后加入phi29连接子和脂质体再水合缓冲液(脂质体缓冲液:3mkcl,250mm蔗糖,5mmhepes,ph7.4)并涡旋。为了得到同质的蛋白脂质体,使用挤出机(avantipolarlipids)将制备的phi29蛋白脂质体通过0.4μm聚碳酸酯膜过滤20-30次。
phi29蛋白脂质体插入minion流动池的共聚物膜中:
minion、没有预插入的生物孔的空白minion流动池和流动池缓冲液从oxfordnanoporetechnologiesltd获得。minion与计算机连接,并且将没有生物纳米孔的空白流动池插入minion中。由oxfordnanoporetechnologiesltd开发和提供的minknow软件,用于按照提供的说明书进行孔插入和数据收集。简言之,将20μl[连接子浓度:200-500μg/ml]制备的蛋白脂质体与280μl流动池缓冲液(25mm磷酸钾,150mm亚铁氰化钾,150mm铁氰化钾,ph8)混合,并上样到灌注口中。使用minknow来施加电压曲线以促进phi29通道插入,随后评估插入了phi29连接子的通道的数量。所有数据通过oxfordnanoporetechnologiesltd提供的定制程序查看。
肽移位:
肽通过genscript合成和纯化。本研究中使用的肽的序列是:
(1)tat:cygrkkrrqrrr(seqidno:1);
(2)mar:markrsntyrsinec(seqidno:2);
(3)r14:rrrrrrrrrrrrrr(seqidno:3);
(4)p27:rrrrrrgfrrrrrrrgfrrrrrrrrrr(seqidno:4)。
将合适浓度(1至20μg/ml)的肽与流动池缓冲液混合,并通过灌注口上样到流动池中。使用minknow观察在不同的施加电势下的肽移位。收集数据后,使用1-2ml流动池缓冲液冲洗流动池,使其可以重复使用。minknow用于证实在将第二肽上样到流动池中之前在minion流动池内部没有肽残留。
电生理学测定:
在teflon分隔膜(孔径:200μm)上形成自立式脂质双层膜。将一对ag/agcl电极置于顺侧-和反侧-腔室中。双层钳位放大器bc-535(warnerinstruments)连接到axondigidata1440a模数转换器(moleculardevices)。在1khz带宽和20khz取样频率下记录数据。使用clampex10(moleculardevices)和clampfit10(moleculardevices)收集和观察数据。
结果和讨论
将phi29dna包装马达的通道插入oxfordnanoporeminion流动池的共聚物膜中:
phi29连接子在脂质膜的电生理学实验中已经被很好地表征(geng等,2010,biomaterials,32,8234-8242;virology,500,285-291)。由于ph29连接子与脂质膜的疏水核之间的疏水不匹配,直接将phi29连接子插入任何疏水膜并不是一件容易的事。因此,通常通过将脂质膜与含有phi29连接子的脂质囊泡融合来进行phi29连接子向脂质膜中的插入。当囊泡和膜由相同材料制成并且分子的疏水尾部具有相同长度时,分子更容易混合。然而,共聚物膜的性质与脂质膜的性质非常不同。共聚物膜的疏水尾部与脂质膜的疏水尾部的长度不同,并且化学性质不同;它们在膜中通常处于更无序和缠结的取向。共聚物的流动性较差且更稳定的性质使得高度有序的脂质囊泡难以与其融合。
令人惊讶的是,该研究的结果已经表明,通过将脂质囊泡与共聚物膜融合,phi29连接子可以从脂质囊泡转移至minion流动池的共聚物膜中。施加的电势也促进phi29通道插入共聚物膜。一旦完成孔插入,插入的phi29通道可以使用minknow通过施加50mv至500mv范围的电压来评估。当施加50mv电势时,插入minion流动池的共聚物膜中的单一phi29通道的电流为约120pa(图5a)。换句话说,在minion流动池缓冲液中,插入minion流动池的共聚物膜中的单一phi29通道具有约2.4ns的电导率(电导率=电流÷电压)(图5a)。为了确认2.4ns电导通道代表插入minion流动池共聚物膜中的单个phi29通道,通过将phi29通道通过脂质囊泡融合插入脂质双层膜中进行了电生理学实验(图5b)。当在oxfordnanoporetechnologiesltd提供的minion流动池缓冲液中进行电生理学实验时,插入脂质膜的phi29通道也显示出约2.4ns的电导率,这证实了在minion流动池中看到的孔确实是phi29连接子蛋白。还观察到多个phi29通道插入minion流动池的单个共聚物膜中(数据未显示)。
已经显示,当施加较高电压(>100mv)时,phi29dna包装马达显示特征性的三步门控行为(geng等,2010,biomaterials,32,8234-8242;virology,500,285-291)。每个门控步骤代表通道大小减少约30%。为了进一步证实minion流动池中产生的电流是由于phi29通道插入,在平台qc过程期间施加-100mv的电压。在-100mv下观察到了特征性的三步门控行为,这表明minion流动池的共聚物膜中的约2.4ns(240pa÷100mv=2.4ns)导电通道代表单个phi29通道(图5c)。
使用插入oxfordnanoporeminion流动池的共聚物膜中的phi29马达通道区分四种肽:
将phi29成功插入minion的共聚物膜中,从而可以对phi29通道进行高通量分析。为了探索phi29通道在minion的潜在生物医学应用,分析了具有不同长度和电荷分布的四种不同的肽。首先,在不同电压下(75mv和100mv)分析12个氨基酸长的tat肽通过phi29通道的移位(图6)。tat肽在两个施加的电势下明显地移位通过通道。在相同的持续时间内,施加更高的电压导致更多的tat移位事件,表明通过phi29通道的肽的传感可以用电压控制。其次,通过插入minion流动池的共聚物中的相同phi29通道,将所有四种肽一个接一个地移位和检测(图7)。在每个肽移位之后和添加下一个肽之前,用缓冲液洗涤流动池。每个单独的肽可以使用振幅和停留时间来区分:tat(图7a)、mar(图7b)、r14(图7c)和p27(图7d)。该实验突出了在生物医学应用中使用单个流动池区分不同肽的高通量能力的潜力。
结论
噬菌体phi29的dna包装马达的通道通过脂质体-聚合物融合插入minion流动池的共聚物膜中。通过观察phi29通道的三步门控验证插入。使用插入的phi29通道区分不同的肽,显示了高通量肽传感的潜力。
除非另有定义,本文所用的所有技术和科学术语具有与所公开的发明所属领域的技术人员通常理解的相同的含义。本文引用的出版物和它们所引用的材料具体引入作为参考。
本领域技术人员将认识到或能够使用不超过常规实验确定本文所述的本发明的具体实施方案的许多等同方案。这些等同物旨在被包括在所附权利要求中。
本文地址:https://www.jishuxx.com/zhuanli/20240726/122243.html
版权声明:本文内容由互联网用户自发贡献,该文观点仅代表作者本人。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如发现本站有涉嫌抄袭侵权/违法违规的内容, 请发送邮件至 YYfuon@163.com 举报,一经查实,本站将立刻删除。
下一篇
返回列表