一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹的制作方法
- 国知局
- 2024-08-02 12:18:19
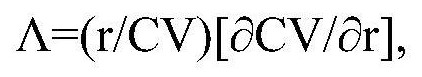
本发明专利涉及细胞生物学、分子生物学、生物光子学、光电子学、激光制导、微分方程。一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹,将细胞受激发光过程的特性光谱信号应用于打开或关闭细胞遗传代谢机制从而加强激光制导导弹技术的信号编码调制,并应用于激发细胞内的激光谐振从而优化激光制导导弹技术的信号增益传输。微分方程数学模型阐释了生物体受激发光过程的光谱特征以及细胞内激光谐振稳定性条件,利于建立质量控制技术指标以促进生物光子智能发射器的设计生产运行。
背景技术:
1、传统的激光制导导弹技术常常面临着“目标信号弱”、“干扰信号强”、“噪音信号多”的挑战。特定场景下对于近距离的敌方军事目标的侦查过程具有如下特点:短探测距离、高探测频率、密集探测方向(目标)、多特征目标和周围多干扰源等,系统并不允许每个探测目标都拥有较长时间的脉冲发射和回波探测。短时间内形成丰富的脉冲编码信息利于提高激光信号的可识别性、抗干扰性。基于受激发生物光子与基因密码的生物激光技术,为激光雷达的信号源发生装置利用生物光子信号带来了新思路。在激光雷达设计过程中通常采用特定波长的红外激光器以及与光源波长匹配的窄带滤光片以提高激光接收系统的信噪比和抗干扰性。多种与激光信号能量相当的干扰信号直接或经反射后被激光雷达系统接收、处理和计算。为保障激光雷达的可靠性与抗干扰性,激光雷达的信号源发生装置亟需提升。光电技术的进步(例如采用光注入、光反馈或光电反馈等方式优化激光光源)利于提高激光雷达的抗干扰性能。新兴的生物光子学(尤其是生物量子光学)为激光制导导弹技术开辟了新的领域,其中的一个优势是生物光子信号的受控激发与基因密码具有紧密关联性,利于实现激光制导导弹系统的超强的“抗干扰信号”性能与“超级密码锁”功能。生物体受激发发光过程中细胞与亚细胞结构的生理与病理过程涉及了复杂的分子遗传学与细胞生物学机制(例如基因密码、蛋白表达与代谢调控)。
2、“生物发光强度”信号在生物光子学研究中被看作是活体细胞的一种特征信号。细胞学研究可以对比分析其它类型的活体细胞信号,比如,磁性纳米粒子标记物的“磁信号”、荧光标记物的“荧光信号”、放射性同位素标记物的“半衰期信号”。萤光素酶不稳定的中间体衰减至基态时发光。萤光素生物发光反应涉及萤光素(底物)、萤光素酶(酶)和分子氧,影响因子包括了阳离子(ca2+与mg2+)以及辅因子(atp、nad(p)h)。
3、与特定生理机制(例如繁殖、喂养与防御)关联的发光蛋白存在于许多海洋腔肠动物中。生物发光蛋白(例如萤光素/萤光素酶系统)一旦与钙结合即可发光。在哺乳动物细胞中表达发光蛋白利于监视应答不同刺激反应的钙变化。生物发光蛋白"指能够发光的腔肠素结合的多肽,"脱辅基发光蛋白"指未与腔肠素结合的发光蛋白。发光蛋白可以通过与腔肠素、分子氧、edta和2-巯基乙醇或二疏苏糖醇温育从脱辅基发光蛋白再生。生物发光(包含发光蛋白)用于实时监测生理或病理机制具有“灵敏度高”、“噪音信号少”等优势(对照荧光系统,它们几乎没有背景)。
4、生物体受激发发光现象具有重要的生物医学应用。基于可见光活体成像原理,带有荧光素酶标记的细胞被应用于实时观测细胞的生理或病理机制(例如,疾病的诱导形成机制、免疫系统中病原转移途径、基因修饰与抗性蛋白表达机制)。活体动物体内光学成像(optical in vivoimaging)采用生物发光(bioluminescence)与荧光(fluorescence)两种技术。生物发光成像技术采用计算机图像软件从选取的活体动物、活体组织、活体细胞及亚细胞结构采集生物光子数据并进行对比分析。通过分子生物学克隆技术以及单克隆细胞技术将荧光素酶的基因稳定整合到细胞的染色体内并培养出能稳定表达荧光素酶蛋白的细胞株。荧光发光是通过激发光激发荧光基团到达高能量状态而后产生发射光。常用的有绿色荧光蛋白、红色荧光蛋白及其它荧光报告基团。利用绿色荧光蛋白和荧光素酶(luciferase)对细胞或动物进行双重标记然后用成熟的荧光成像技术进行体外检测。将分成互补性两段的荧光素酶基因分别连接待研究的两种蛋白之一的编码dna,然后经导入细胞或动物体内被表达为融合蛋白。生物发光被用于观测活体干细胞中两种蛋白质的相互作用(预先让两种蛋白质各自携带荧光素酶的互补性片段结构,存在强相互作用的两种蛋白质彼此靠近,原本被隔离的荧光素酶片段结构在相互靠近与共同作用于底物的过程中展示生物发光特性)。生物发光被用于判别细胞凋亡的进程(荧光素酶与抑制多肽对应的融合蛋白无荧光素酶活性,在特异识别位点切割抑制蛋白利于恢复荧光素酶活性)。
5、已经公开的用于促进激光制导技术的专利涉及了激光编码技术(通过对激光的脉冲宽度、脉冲间隔、脉冲幅值、重复频率、波长及相位等参数进行调制以区别于其它激光信号)。已经公开的关于生物发光的专利涉及了基因修饰、蛋白质表达、高通量筛选、荧光医学检测与干细胞等前沿技术。
6、专利(cn107831486b)使用基于ram方式的脉冲相关处理方法,在fpga内部采用脉冲索引与匹配算法,对高频干扰脉冲可迅速地匹配到正确的目标反射脉冲串。
7、专利(cn102638271a)对于激光制导(跟踪)系统中纳秒级脉宽可调激光器提出一种使用电光开关直接削波实现脉冲宽度和脉冲幅值的编码以对抗激光干扰。
8、专利(cn105182361)提出利用信号发生器产生的复合调制随机脉冲信号将脉冲位置调制和脉冲幅度调制相结合的一种脉冲编码序列的调制方式。
9、专利(cn112817335a)公开了公开了一种激光驾束制导飞行器的制导控制方法,采用多次幂趋近律滑模控制的分段制导方式完成制导飞行器整个飞行过程的制导律设计,实时解算舵偏指令和制导指令并据此控制飞行器沿激光方向命中目标。
10、专利(cn107885230b)公开了带有激光制导武器的无人直升机控制系统及控制方法,所述机载子系统包括机载数据链终端、稳瞄装置、飞控模块、武器模块和机载任务管理器(武器上设置有含有激光接收器的激光导引头)。
11、专利(cn101370823a)公开了“发光干细胞及其应用”,描述了能在生色团底物存在下响应于细胞内钙浓度变化而表达脱辅基发光蛋白且生成生物发光信号的稳定的重组干细胞。
12、专利(cn1317299c)公开了具有较高的发光活性的嵌合发光蛋白质(可用于检测胞内钙),其制备方法是通过将奥贝林蛋白质所包含的位于第一个和第二个钙结合位点之间的区域用选自clytin、水母发光蛋白、thalassicolin、mitocromin、mnemiopsin和berovin的发光蛋白质中的相应区域进行替代而获得。
13、专利(cn1997383a)提供了含水母发光蛋白的组合物,以及它们用于预防和/或减轻与钙失调相关的症状和障碍的方法。
14、专利(cn104781401a)基于数据库中荧光素酶氨基酸序列人工构建一系列氨基酸序列并提供一种高亮度且发光稳定性高、向长波长侧迁移的光谱。
15、专利(cn108796041b)公开了一种基于生物发光共振能量转移的信号扩增系统(包括:能与dna结合的能量供体蛋白质;能与dna结合的能量受体蛋白质;由三种dna探针所组成的核酸组装体系)。
16、专利(cn101634634b)利用金属纳米线中传导的表面等离子体将激发光传播到细胞内,实现对细胞内物质或标记物的纳米范围的远程激发。
17、专利(cn112557353b)公开了一种基于延迟发光光谱的细胞活力检测方法和装置(在检测细胞延迟发光的装置基础上增加不同波长的滤光片,短波长滤波片的波段范围是300nm~500nm,长波长滤波片的波段范围是500nm~650nm)。
18、专利(cn1712939a)公开了一种基于激光诱导荧光的细胞成像方法。标记染料的细胞在激光照射下发出荧光由显微镜物镜收集,经滤光片在ccd监测器上成像并得到细胞的荧光照片。
19、然而,这些专利存在以下问题和缺点:这些专利没有将生物体受激发光技术应用于激光制导导弹。原因之一是两个学科领域差异大、技术门槛高;原因之二是我国整体科研创新能力落后于欧美发达国家(制约性因素包括了专家团队的组建、研发与产业化配套资金、关键设备的研制与核心技术经验的积累、项目评审验收与质量管理体系认证能力)。
技术实现思路
1、为了解决上述问题,本发明提供了“一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹”,所述生物体受激发光技术涉及核酸遗传、基因密码、蛋白表达、代谢调控等一系列分子遗传与细胞发光生物学复杂机制。
2、一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹,其特征之一在于,该过程包含如下步骤:“受激发的生物体发光”-“生物光子信号采集”-“生物光子信号编码调制”-“生物光子信号传输”-生物光子信号解调识别”-“激光制导”;所述激光制导导弹技术的“生物光子信号编码调制”、“生物光子信号传输”与“生物光子信号解调识别”分别对应着“生物光子信号发射器”(由光敏物质靶向性激活的生物发光蛋白,包括但不限于采用了“脱辅基发光蛋白”、“嵌合荧光基因片段”、“荧光素酶基因标记”等方法)、“生物光子信号传输装置”、“生物光子信号处理器”。
3、“生物光子信号发射器”源自生物体发光细胞中发光蛋白或纳米尺度的激光谐振器。借助计算机软件系统把连续性的军事目标信号转换为生物体的发光细胞的入射光信号,“生物光子信号发射器”能够转换细胞的遗传代谢机制中“分子关闭”与“分子激活”工作状态并高效率地将军事侦查目标对应着的能量层级差异性很小的连续性信号区分为易于编码调制的阶跃性数字信号。探索生物体发光细胞的受激发光过程的动力学特性以及纳米尺度的激光谐振器的稳定性条件,将突破性地把连续性信号转化为阶跃性的数字信号,利于增强激光制导导弹系统的信号编码调制与信号解调识别能力,尤其是利于开发适用于激光制导导弹系统的具有“信号保真性能高、抗信号干扰能力强”特点的源自生物体发光过程的信号源。
4、“生物光子信号发射器”的特点是“发光生物体处于受激发发光状态”、“生物体受激发光过程的内在调控机制涉及基因遗传密码锁”、“采集并处理了受激发的生物光子信号”。“细胞发光生物学的不确定性”涵盖了细胞的物质代谢、能量代谢与基因遗传等方面的“黑匣子效应”。质量管理体系利于监管所述生物发光强度信号的各个环节(例如发光细胞将具备“细胞物质代谢活性高”、“细胞能量代谢活性高”、“细胞发光活性高”等优点),从而确保从实验室阶段迈入到“规模化”、“规范化”与“标准化”的产业阶段。
5、“生物光子处于受激发状态”:生物光子普遍性地存在于细胞的生理或病理过程中。广义的生物光子可以被定义为活体生物、活体细胞与亚细胞结构产生的光子信号,这些信号可以被高灵敏度的仪器设备采集并进一步作数据分析处理。生物光子(微量或微弱的生物荧光信号)可以是由单一的具有特定结构的蛋白质所产生,也可以是由涉及多种蛋白质协同作用的某种细胞代谢通路或细胞信号传输通路所产生。
6、“生物光子信号发射器”由光敏物质制导并激活生物发光蛋白,用于靶向制导地精准有效地激发生物光子。促进生物发光的方式包括但不限于采用了“脱辅基发光蛋白”、“嵌合荧光基因片段”、“荧光素酶基因标记”等方法。涉及发光蛋白的基因修饰与分子遗传方法如下:用本领域技术人员已知的标准转染方法,包括但不限于,电穿孔、peg ca沉淀、阳离子脂质方法等,可以实现稳定重组。有利地,稳定的重组干细胞可以分化成特定细胞语系,以表达至少一种特定细胞谱系靶物。脱辅基发光蛋白意指任意的脱辅基发光蛋白(天然的或重组的或合成的)。脱辅基发光蛋白具有提高的发光活性和/或钙灵敏度,其来源可以是天然的或诱变的突变体(例如源自不同的天然脱辅基发光蛋白的嵌合蛋白)。为维持或增加发光蛋白在光发射和钙响应性方面的活性,可以通过选择密码子序列、融合至线粒体靶序列、或删除、添加或置换氨基酸残基从而实现优化脱辅基发光蛋白。
7、作为非限制性例子,下边阐述基于atp酶的“生物光子智能发射器”以及对应的生物发光强度信号机制:
8、生物光子链式反应是受激发的生物光子产生过程。在正常生理或病理机制中,发光蛋白产生的生物荧光可以被高精度的仪器设备采集并进行进一步的数据分析处理。“受激发”指发光蛋白产生生物荧光的过程得到加强或促进。atp酶(分子马达)中发光蛋白产生生物荧光的过程,可以通过增加发光蛋白的表达量或增强发光蛋白的发光能力等方法让发光蛋白处于“受激发”状态。其中,增加发光蛋白的表达量对应着基因表达与调控过程,增强发光蛋白的发光能力对应着采用合适的小分子或大分子维护发光蛋白的活性中心。在输入氨基酸的条件下调控细胞的生理或病理机制中普遍存在的三羧酸循环(atp是呼吸链的关键性环节之一)。下边举例介绍了具有增强的生物发光性能的发光蛋白:借助特异性的或可诱导的启动子可以克隆脱辅基发光蛋白编码序列,然后通过嵌合目标蛋白的方式可以得到发光蛋白。例如在下述位置gly142-cys诱变的clytin序列(genbank登记号q08121):ala9-gln,val92-leu,glu97-gln,ala-cys,ile78-cys,val86-lie,glu87-ala。
9、作为非限制性例子,测试生物发光强度信号的步骤如下:用腔肠素在室温温育3h,测定每个组织中的发光蛋白含量。注射250mm cacl2和triton x-100溶液。基于ccd照相机的光度计条件高灵敏度,读数时间60秒。利用通过先进的分子影像监测设备pet/mr对细胞内生物光子产生位置准确定位,再利用高能电子加速器la45进行高能(45mv)与低剂量(4-5gy)x线照射,细胞内产生的双氧水将与光、光敏剂产生大量的自由基和单线态氧。低剂量高能光子、光敏物质和催化底物均无明显毒性,但三者作用产生的产物(自由基和单线态氧)具有强大的细胞免疫保护作用与异体组织清除作用。
10、2一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹,其特征之二在于,所述“生物光子链式反应”的运行机制具有“微观复杂性”、“微观敏感性”的一种链式反应,所述“纳米尺度的能量转换器”具有“实用性”与“高效性”特点。
11、分子开关指基于光或温度的变化或通过结合溶液中特定的离子或分子来实现开/关。光信号或外加的电磁场作用于光敏型的分子开关从而调控发光蛋白的生物发光过程。工作模式在开启/关闭两种状态之间转换。类似的作用机制(分子在“关闭”与“激活”状态之间进行转换)利于实现超微结构内(例如10微米尺寸的生物体组织)的靶向控制。发光蛋白的表面结合了一个光敏型的分子开关利于控制发光蛋白的运动过程。一个非限制性例子是,分子内含有一个氮氮双键对应着分子处于“关闭”状态,当用蓝光照射分子并打破氮氮双键之后分子由“关闭”转为“激活”状态(由双键连接的两部分会发生旋转从而激活分子)。
12、受激发生物光子由细胞或亚细胞结构中“纳米尺度的能量转换器”不同于量子物理的隧穿过程。原子核裂变或聚变对应的链式反应可以被激发。核反应堆的调控措施被简单地阐述为“升降控制棒”从而减缓或提升核反应堆的链式反应速率。发光蛋白(由原子组成)可以被看作是纳米尺度的能量收集器,其所需驱动力可以来自热能或电能(例如,在室温下以每秒几百万转的速度随机旋转)。由电子扫描显微镜产生的电能会导致方向旋转(电能从其尖端流入发动机,单电子的能量足以使转子继续旋转仅仅六分之一圈)。
13、下边将生物光子链式反应与原子核裂变或聚变对应的链式反应进行比较分析:
14、链式反应都对应着微观复杂性:生物光子链式反应存在于细胞或亚细胞结构中,与一系列的生理或病理机制相关联;原子核裂变或聚变对应的链式反应对应着一系列复杂的机制(涉及到微观的物质结构改变与巨大的能量层级改变过程)。
15、链式反应都对应着微观敏感性:生物光子链式反应存在于细胞或亚细胞结构中,受制于一系列复杂的细胞信号机制、能量代谢机制、物质传递机制;其“微观敏感性”体现在“微小的变化,来自于细胞信号、能量代谢或物质传递的任何一个因素,都可能引起生物光子链式反应产生差异”。原子核裂变或聚变对应的链式反应受制于一系列复杂的中子或质子产生机制、中子或质子捕获机制、原子核裂变或聚变机制;其“微观敏感性”体现在“微小的变化,包含来自于中子或质子产生或被捕获、原子核受激发而产生裂变或聚变中的任何一个因素,都可能引起原子核裂变或聚变对应的链式反应产生差异”。
16、“纳米尺度的能量转换器”具有“实用性”与“高效性”特点:
17、能量供应与细胞工作机制的转换之间存在着微妙关系(例如,细胞膜上离子通道对于外界的物理、化学或生物信号作出信号反馈)。位点结合的相互作用是带正电区域与带负电区域之间的静电吸引作用(并不是高强度的共价键)。相对较弱的结合力让聚集方式可以被打破或重建(类似于双链dna间的氢键)。分子首先需要克服哑铃型链状分子一端的空间壁垒,与一个可切换的连接位点相结合。之后连接位点改变自己的结合状态,跨过第二道壁垒从而到达链状分子的存储端。
18、细胞中发光蛋白利用离子或是分子的浓度梯度差异来调控生理或病理过程中蛋白酶的催化进程。体外测试过程中发光蛋白的表观运动速度(每秒行进的距离)是自身长度的1,000多倍。发光蛋白的表观运动速度并不高,却在生物光子信号产生过程中体现了极高的生物活性。形象性的表述方式如下:核糖体高效率地组装氨基酸分子,蛋白酶在运动过程中捡拾氨基酸分子并添加到一个不断增长的肽链上(经过大致估算,1018个这样的分子流水线在超过36个小时的时间里合成出了几毫克的多肽)。
19、3一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹,其特征之三在于,所述激光制导导弹技术中受激发生物体发光过程采用了质量管理体系,对于具体实施方案进行了质量控制与管理(涵盖了工艺技术指标的拟定以及质量指标的检测与验收)。有益效果是,利于将所述生物发光强度信号的各个环节纳入到质量管理流程,促使本发明的技术方案从实验室阶段成功地迈入到具有“规模化”、“规范化”与“标准化”的产业阶段。生物光子信号具有“微弱性”、“分散性”、“灵敏性”特点,在信号传导过程中并不能替代激光制导导弹的激光束。但是,通过“生物信号加密编码”可以把生物发光强度信号加载到激光束,对应着一个具有深远意义的“信号加密”应用领域。
20、所述“受激发生物光子的基因遗传机制”受制于“基因遗传密码”、“细胞代谢调控”等复杂的生理病理机制,对应的“生物信号保密编码”过程具有“复杂性”、“规律性”、“特异性”等特点。基于“基因遗传密码”与“细胞代谢调控”等生理病理机制,本发明证实了“生物信号保密编码”的“可行性”与“实用性”,利于“激光制导导弹系统”具备“信号保真”与“信号加密”等优异性能。具体而言,采用质量管理体系监控细胞的制备过程从而提升细胞的生物发光指标(具备“细胞物质代谢活性高”、“细胞能量代谢活性高”、“细胞发光活性高”等优点)。
21、从实用性角度,本发明阐释了信号收集、信号加载、信号传输、信号解调过程中生物发光强度信号的作用机制、调控机制,从而将细胞基因遗传密码与激光制导导弹系统这两个差异悬殊的学科领域衔接起来。“生物信号加密编码”是将微弱的生物发光强度信号加载到激光束。该过程类似于无线电信号传输过程中的“信号调制”,即把低能量的信号加载到高能量的传输载体。后续的“解调”过程可以取出“信号调制”中加载的生物发光强度信号。从细胞基因遗传密码角度,细胞的生理病理过程中能量代谢途径或物质代谢途径激发了生物发光,对应的调控机制是细胞的生理病理过程受制于内部的细胞因子或外部的环境因子。本发明给出的实施方案利于将模拟信号转变为数字信号,从而利于将复杂的外界军事信息由模拟信号转变为数字信号。激活过程对应着临界值(低于临界值,不会被激活;超出临界值,出现激活现象)。通过信号处理装置设置输出信号的临界值,利于实施“启动开关”的控制作用。启动开关的开启状态对应着二进制信号中“1”,启动开关的关闭状态对应着二进制信号中“0”,这样,产生了数字信号。这样,将输入的模拟信号,通过细胞的生理或病理机制,转换成数字信号。有益效果是,外界的模拟信号被转变为数字信号,利于实现信号传输过程的远距离传输、高保真传输与高保密传输。
22、本发明的技术策略是调控光刺激以控制活体组织中的细胞(尤其是神经元细胞)。生物光子信号需要放大,然后聚集效应或经过放大的生物发光强度信号被加载到激光束。作为一个放大生物光子信号的实施案例,运算放大器为采用单电源供电、线性稳压器、闭环pid控制的低温漂型运算放大器(温度变化<0.5℃,生物荧光检测器工作时功耗<10.7mw)。从分析检测设备方法看,采用高通量筛选(hts)优选发光蛋白的生物活性,采用生物发光共振能量转移(bret)研究蛋白相互作用;从细胞的制备过程看,病毒(附有发光感应器)通过感染神经元从而进入了神经元细胞内部;从细胞活性的调控机制看,基于发光蛋白的发光特性(钙离子激活腔肠素/水母蛋白复合物产生生物发光强度信号),选择钙离子作为神经元活动的信号标志。本发明的实施方案采用激光照亮细胞信号通路。低于阈值的输入信号对应着微弱的输出信号,但高于阈值的输入信号可能使输出信号变得巨大而且呈现非线性变化趋势。被安置在移动式飞机上的激光照射机对应着很高的运营成本(飞机飞行对应着机油损耗、机械折旧等常规消耗、人工成本、意外伤害)。固定位置的激光照射机在对欲打击目标进行照射后,激光制导导弹的引导头立即接收到了回射的激光。从信号接收角度看,激光制导导弹的激光信号可以来自某个固定位置的激光发射器(比如悬崖绝壁的崖石尖端)。
23、作为非限制性例子,以发光传感器探测大规模神经元的实验步骤如下:把发光感应器插入老鼠海马体的大脑切片(含有数千个神经元)中,以较高浓度的钾离子打开细胞的离子通道。发光传感器通过增亮和变暗来响应钙浓度的变化。神经元存在“兴奋性”与“抑制性”机制(人脑是由大约1000亿个神经元组成,其中每一个神经元通过数千计的微小突触接收来自其它神经元的电信号。当信号在其突触的总和超过一定的阈值时,神经元便会发光)。
24、下边以钙离子激活发光感应器作为例子表明了质量管理体系贯穿于发光感应器验证环节:钙离子激活发光感应器的机制:发光感应器一旦遇到钙离子就会发光;其次,钙离子参与神经元的活化过程(神经元外的周围环境中钙含量往往较高,但是细胞内部含量很低,在神经元受到来自相邻的另一个神经元的刺激时,钙含量会短暂达到尖峰水平)。由腔肠素、脱辅基水母发光蛋白(apoaequorin,apo)和分子氧反应生成复合物形成活化的水母发光蛋白(aequorin),一旦与钙离子结合后,复合物结构被破坏,腔肠素被氧化生成高能产物(coelenteramide),同时释放出co2和蓝色光(~466nm)。
25、实验室结果表明:阻遏蛋白和g蛋白在调节gpcr功能过程中分别扮演阴和阳的角色(gpcr能激活g蛋白的信号通路,而阻遏蛋白会识别被激活的gpcr并内吞到细胞内进而阻止g蛋白向下游传递信号)。视紫红质复合物结构中存在90°旋转的构象差异(受体可能处于n端结构域的前面方位);视紫红质gpcr可以感应到光信号;g蛋白偶联受体(gpcr)将信息通过下游g蛋白发送到细胞;g蛋白和阻遏蛋白构成了gpcr下游的两条主要信号通路。非视觉阻遏蛋白参与调控多个gpcr下游信号通路(不仅阻断受体与g蛋白的结合,还介导受体的内吞及一系列非g蛋白依赖的信号通路)。非视觉阻遏蛋白与gpcr的相互作用较弱(组装的复合物具有高度的柔性)。视紫红质感受外界光信号并将光信号传导到细胞内产生视觉。基于视觉细胞仿生学原理,考虑到视觉细胞对于光源有高效率的光敏性,重点开发视觉细胞中视紫红质—阻遏蛋白复合物晶体结构的生物医学应用。
26、嵌合光敏蛋白的动物学实验步骤包括如下内容:基于立体定位法确定目标位置后注射携带光敏蛋白的aav(病毒体积需根据病毒滴度和核团大小等调整),待病毒表达2-3w后,取小鼠断头取脑,震荡切片,片厚300μm(目标脑区)。需要进行比较分析的环节如下:追踪脑区中被aav荧光标记的神经纤维附近状态好的细胞,比较细胞响应与光刺激之间的时间间隔。当细胞的信号反馈能力不符合预期,需要判别制约性环节或负面因素。例如,在确信细胞状态良好的前提下通过增加刺激强度来判别病毒转染过程中载体是否具备活性。
27、实验室研究表明:在信号输入层面,以存在间隙时间的钛蓝宝石激光脉冲信号作为输入信号(当受到两个输入脉冲间隔小于150ps时),能够检测到一次生物发光强度信号。用800纳米的钛-蓝宝石激光器激发纳米二极管激光器阵列,结果显示某些几十纳焦的脉冲信号在200皮秒时间范围内可以激发生物发光强度信号。接受外部刺激后细胞以生物发光强度信号作出信号反馈的时间间隔是毫秒级的短暂时间。当两个脉冲发生的时间间隔为150和350ps时,细胞具有“相对的不响应时期”(由细胞信号反馈机制产生的生物发光强度信号会比较弱)。此情况下细胞的生物发光强度信号只对应于一个初始脉冲信号。
28、测试受激发生物光子信号发射器效率的试验方案:技术核心位于激光照射器模块中生物光子的受激发过程、生物光子信号的加载(生物光子信号被纳入激光信号的编码过程)。试验方案包括激光制导导弹模块、激光照射器模块、激光传输效应模块和导弹目标;激光制导导弹模块的导引头根据反射激光信号引导激光制导导弹模块的导弹调整航道和追踪导弹目标;激光传输效应模块用于提供不同天气条件下入射激光和反射激光传输时的能量衰减数据。导弹目标的工作模式能够模拟导弹目标:坦克,能够设置坦克的型号和表面漫反射率。将单个脉冲周期内的激光信号转变为多个强度可调制的子波(具有特征子脉冲数,特征脉宽,特征时间间隔、特征强度顺序)。基于子波的脉宽、时间间隔和激光强度顺序实现激光信号编码。激光雷达的回波信号处理系统筛选回波信号。
29、4一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹,其特征之四在于,采用了激光信号发射装置、生物光子信号发生装置、生物光子信号的激光编码装置、激光制导导弹的信号接收装置等设施设备。
30、生物光子链式反应对应着“纳米级原子弹”:细胞大量吸收光敏药物后定向于细胞线粒体atp酶,激发的生物光子链式反应类似于纳米级原子弹。激发产生自身特异性免疫应答并产生生物光子的过程类似于原子弹冲击波得到扩散;细胞自身特异性免疫应答机制保证了细胞代谢层面的“安全无毒”与人工操作层面的“精准高效”。信号源存在着“能量损耗低”、“信号保真高”、“信号干扰少”等优势。其中,“能量损耗低”优势利于降低信号发射塔的运营成本;“信号保真高”的优势利于维护信号发射塔的军事机密性;“信号干扰少”的优势利于对抗潜在的军事对手实施的信号干预手段,并利于增强军事攻击时激光制导导弹系统的精准定位能力。
31、可以将生物光子信号发射器设置在人迹罕至、有毒物质风险区、噪音污染风险区、放射性污染风险区、过高或过低的温度与湿度区域、高纬度或高海拔区域等位置,然后基于生物光子信号发射器的“实时监测”、“灵敏度高”、“准确度高”等特点,把敌方军事目标信息反馈在生物光子信号的编码调制与解调识别过程中。
32、信号采集装置可以采用基于激光诱导荧光的活体细胞实时检测方法及装置(包括光路、与光路连接的信号处理系统)。光路包括激光发射光路、散射光接收光路和荧光接收光路。生物光子信号发射装置可以增强放大微弱的生物光子信号并发射激光信号,借助计算机软件系统进行编码调制与解调识别,然后应用于激光制导导弹系统。
33、作为非限制性例子,下边介绍了高性能的生物光子信号的产生装置与信号传输装置。
34、表达发光蛋白的干细胞或其分化的细胞经过信号采集与信号反馈过程可以产生生物发光强度信号。基于发光蛋白的测定与所有钙敏感的荧光染料的高荧光背景信号相比,几乎没有背景,体现在发光响应相对于荧光响应的更大的线性动态范围。激发光波长可以是460,470,490,515,530,560,590,640nm;光敏通道蛋白包含光激活与光抑制两种类型。三磷酸腺苷生物光子发生器可以被用作生物光子信号发生器。细胞大量吸收的光敏物质可以穿过线粒体膜参与细胞呼吸链atp催化过程;经pet-ct/pet-mr定位细胞中atp蛋白复合体,再进行高能加速器照射。光敏物质(例如经改良的卟啉)具有对细胞的“高特异性与高亲和力”结合作用以及分子靶向作用等特点(高亲合力体现在细胞的特定结构位置的浓度比细胞其它位置浓度超出10-20倍;分子靶向能力体现在可以精准结合到细胞内的线粒体蛋白复合物)。细胞内三磷酸腺苷(adenosinetriphosphate,atp)含量的变化可以间接反映细胞杀伤、细胞抑制和细胞增殖过程中细胞活性(在0.5~5mmol/l的浓度范围内具有良好的线性关系,相关系数为0.98,最低检出限为0.06mmol/l)。
35、光敏感通道蛋白(channelrhodopsin-2,chr2)是一种受光脉冲控制的具有7次跨膜结构的非选择性阳离子通道蛋白。光激活蛋白包括了cheta与hchr2,光抑制蛋白包括enphr3.0与hgtacr1。在1-10um生理范围内的发光蛋白的钙高灵敏度,允许甚至在小量钙运动情况下检测。发光蛋白具有靶向特定细胞隔室如线粒体的可能性,以便以更精确的方式检测从内部储存的钙释放。:
36、下边给出了应用光敏感通道蛋白的生物发光强度信号的信号产生与信号采集过程。光敏感通道蛋白涵盖了兴奋性与抑制性光敏感通道两种类型。
37、关于生物光子信号发射器的通用性实施方案如下:a)将某种细胞分化成特定细胞谱系;b)给细胞装载合适的生色团作为底物;c)使细胞接触化合物文库(合适的光敏感通道蛋白插入病毒载体中并感染细胞,使光敏感通道蛋白在特定类型的细胞中表达);d)检测发光蛋白的生物发光(通过特定频率的激光调控光敏感通道蛋白)。e)寻找合适的充当报告基因的视蛋白(包括br、hr、chr、cochr等);向受体细胞输送光敏感蛋白,完成转染和表达;f)改变光照波长和光脉冲强度以调控生物光子信号。作用机制如下:蓝光照射让chr通道开放并引致阳离子大量内流,该去极化过程诱发的动作电位可以激发细胞产生生物发光强度信号。黄光照射让hr通道开放并引致阴离子大量内流,该超极化过程抑制了动作电位从而阻碍细胞产生生物光子信号。
38、关于生物光子信号发射器的一个概念化实施案例如下:集成电路带有光传感器、发送器和微电路芯片(检测对象可以是核酸、蛋白质、细胞、微生物等)。寄存器芯片组将控制指令的串行输入转换为并行输出(控制发光二极管的亮灯时长、闪烁方式、灯光亮度以及光色);处理器寄存器芯片组与发光二极管寄存器芯片组相连。蓝牙无线通讯主机通过usb接口与上位机pc相连。利于高通量检测的设计方案如下:发光二极管组包含96个发光二极管,成8×12矩形排列,各发光二极管间中心距离为9mm,与生物细胞实验所用标准96孔细胞培养板各孔位置相对应。芯片可以将检测结果传送到外部的计算机软件系统,借助激光传送过程把生物发光强度信号用于激光制导导弹的信号编码调制从而实现远程控制。生物光子信号发射器包括支架、控制开关和两个玻璃容器(其中一个玻璃容器是生物光子链式反应的场所,一个玻璃容器被用来储存营养成分)。两个玻璃容器通过控制开关相互连通以控制荧光素的流速。发光二极管的出射光波长包含470nm、520nm以及620nm,工作频率为40~100hz,以pwm方式调节工作亮度并对细胞实施光刺激。
39、5一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹,其特征之五在于,受激发生物光子可以为激光制导导弹技术提供基于基因遗传机制的激光信号密码调制方法。本文采用数学物理模型探讨了生物光子智能发射器中关于受激发生物光子的信号产生、信号衰减或增强过程。
40、基于基因突变手段获得了一系列光敏感通道蛋白,对应的时间特性和激发光要求具有多样化特点;多样化的光敏感通道蛋白能够满足特定用途的生物发光强度信号的个性化定制需求。与之配套的检测“受激发生物光子”强度的方法包括但不限于采用“基因修饰与蛋白表达”技术、“生物光子采集”技术、“生物发光定量分析”技术。采用激光激发生物发光强度信号具有“高时空分辨率”、“高特异性”以及“高选择性”等特点(调控激光可以将时间精准到毫秒级别甚至是亚毫秒级;将光敏感蛋白锚定在靶向细胞或细胞器的措施包括但不限于定位注射、特异性启动子以及亚细胞器定位肽)。
41、5.1建立数理模型的目标任务
42、5.1.1信号转换:本文通过数学物理模型论证了“可行性”与“实用性”。对于细胞的信号感知过程与信号反馈过程,外部环境影响因子包含着多种可能的刺激因素,例如,光源刺激、热源刺激、电场刺激、磁场刺激、声源刺激、化学或生物来源物质的刺激、甚至是核辐射刺激。细胞结构具有“生命特征性”、“感应灵敏性”、“条件温和性”。为了避免外部环境影响因子损伤细胞,预先对外部环境影响因子进行合规化处理。例如,“合规性”信号处理的例子是,通过信号过滤去除干扰信号或破坏性与损伤性信号(过高强度或过高频率的刺激信号)。通过转换细胞的遗传代谢机制中“分子关闭”与“分子激活”工作状态,将军事侦查目标对应着的许许多多能量层级差异性很小的连续性信号区分为许许多多的易于编码调制的阶跃性数字信号。本发明所述“潜在的敌方军事力量或敌方军事活动”对应的军事信号,可以产生上述刺激因素,或者是可以被转换为上述刺激因素,从而形成被细胞感知的外部环境影响因子。
43、事实上,可以通过合适的信号采集装置与信号转换装置,把外部军事信息中的其他情形转换为本文提及的上述两种常见情形。从理论上来说,任何外部军事信息都直接或间接地与上述两种常见情形(光信号与电磁信号)存在着关联现象(事物之间存在着普遍联系性)。参照上述两种常见情形(光信号与电磁信号),其它的外部军事信息也可以经历被采集与被转换过程,从而进入到后续的细胞接收信号与细胞反馈信号过程。当然,外部军事信息的特异性与复杂性等特性也意味着对应的信号采集与信号转换装置具有特异性与复杂性等特性。
44、5.1.2信号输入:考虑外部军事信息中的两种常见情形(光信号与电磁信号),本发明采用一个信号采集装置与信号处理装置,将获取的光信号与电磁信号转化为一种输出信号,比如说某种光信号。这种光信号将被细胞所感知并被细胞所反馈。光遗传学(optogenetics)指结合光学与遗传学手段精确控制特定神经元活动的技术。利用基因操作技术手段将外源光敏感蛋白基因导入活细胞,在细胞膜结构上进行光敏感通道蛋白的表达。然后通过特定波长光的照射,完成细胞膜结构上的光敏感通道蛋白的激活与关闭,以控制离子通道的打开与关闭,进而改变细胞膜电压的变化,如膜的去极化与超极化。当膜电压去极化超过一定阈值时就会诱发神经元产生可传导的电信号,即神经元的激活。相反,当膜电压超极化到一定水平时,就会抑制神经元动作电位的产生,即神经元的抑制。
45、5.1.3信号处理:把细胞作为灵敏的信号接收装置与信号反馈装置,需要预先把外部环境因子进行信号处理(signal processing),该过程的作用是把外部环境因子信号由“不规则性”或“不确定性”转换为“合规性”与“确定性”,从而利于被细胞感知并反馈。
46、下边阐释了生物光子信号技术的预备阶段(信号采集与信号转换过程)以及生物光子信号技术的实施阶段(细胞感知信号并反馈信号的过程)。生物光子信号技术的预备阶段(信号采集与信号转换过程):宏观世界(比如潜在的敌方军事力量开展的军事侦查甚至军事入侵活动)存在着信号产生与信号传输过程,相应的的外部军事信息可以被收集。为简化表述,下边仅仅列出外部军事信息中的两种常见情形(光信号与电磁信号)。
47、5.1.4关键任务:本文侧重讨论激光制导导弹的信号加载、信号编码与调制阶段,尤其是生物发光强度信号的产生阶段。本发明的意图是挖掘生物发光过程受生物学密码制约的内在规律性,指明生物体受激发生物发光过程具有“密码特异性”、“信号保真性”、“传输高效性”等特点,有利于将生物体受激发生物发光过程嵌入信号编码与信号传输过程从而大幅度提升激光制导导弹系统的“信号采集与调制传输”性能。生物体受激发生物发光过程受制于基因遗传与代谢调控机制中的生物学密码(基因核苷酸密码与氨基酸序列密码),输出的信号是生物发光强度信号。基于生物体受激发生物发光过程与生物学基因遗传密码并不存在严格意义上的一一对应关系的客观事实,本发明并不能直接破译生物学基因遗传密码,也不能将生物学基因遗传密码直接地融合于生物体受激发生物发光过程。本文的目的不是探讨激光制导导弹的运动过程(该运动过程中空气阻力不可以被忽略,对应的加速度是复杂多变的变量)。
48、5.1.5任务分解:本发明建议将数理模型研究进行任务分解。本发明建议将生物发光过程的细胞参数方程分解为多个子模型(其中每个子模型只考虑生物发光过程的一个子集,然后对每个子模型进行近似求解),最后将结果集成得到完整模型的解决方案。通常关于生物发光过程的细胞参数的主方程难以获得对应的精确解。为简化讨论,假定在时间t内的细胞发光能力λ(cell glowing ability parameter)是一个恒定的数值,意味着现实世界中生物体(发光细胞、细胞、病毒等)的内在的发光能力遵循复杂的变化规律,但本文的简化讨论将复杂变化的数值简化为均匀恒定的数值。内在的发光能力制约着生物发光强度信号产生变化的速度,大体上对应于牛顿第二定律中的加速度(速度在单位时间内的改变量)。它由生物体发光的内在因素(基因遗传与代谢调控机制)所制约并成为了生物发光强度信号的内在推动力。初等物理学中将多样化的加速运动简化为牛顿第二定律中具有恒定加速度的情形(例如忽略空气阻力的情况下自由落体运动中物体在恒定的重力加速度作用下撞击地面)。
49、5.1.6实施策略:本发明采用数理模型论证了生物发光强度信号可以被用于数据编码与数据传输过程。1需要配备高精度检测设备:发光细胞(或发光细菌、发光病毒、发光藻等)受激发后的生物发光强度信号是间断性的微弱光信号。因此对应的光信号采集装置需要具备“高灵敏度”与“相应速度快”等特征。2参数调节与数据处理:实证研究需要测试的数据为生物体的光谱特性数据(光谱形状、峰高与半波宽度等)、恢复期。考虑到峰值波长是特定的生物体固有的属性,在生物发光发射装置的功能调试环节可以任意改变输入信号的强弱,采用由多次测量结果中取平均值的方法来确定峰值波长。
50、5.2对于实验研究结果的概述
51、5.2.1生物发光强度信号的特异性:考察了多个因素影响(信号输入条件、生物体生长条件中浓度温度与酸碱度、生物体生长期)之后,生物体具有特定的光谱特性。发光细胞(或发光细菌、发光病毒、发光藻等)受激发后的生物发光强度信号具有类似于“指纹图谱”的特异性(光谱的基本形状以及峰值位置)。不同生物体之间的发光能力存在明显差异。例如,在月芽形梨甲藻、夜光虫、多边溪沟藻之间,月芽形梨甲藻的闪光最亮、夜光虫的闪光频率最高、多边溪沟藻的闪光亮度最弱。此差异性根源于细胞的类型、尺寸、闪光机制的能量与物质供应能力等因素。以发光藻为例,生物发光是细胞内生物化学反应导致电子跃迁的结果,其内在的运行机制受制于生物体固有的基因遗传与代谢调控机制,不因外部的信号刺激发生偏移或改变。发光藻的峰值波长约为472nm,谱线的半波宽度(fwhm)约为30nm。
52、5.2.2生物体细胞的持续发光能力:第一次发光强度会明显超过持续状态的发光强度;生物体在间隔一段时间(比如说,恢复期为六个小时)后恢复最佳生理状态并展现初始入射光刺激条件下的高峰值发光能力。两次入射光刺激的间隔时间过短(例如小于恢复期的三分之一),生物发光强度信号可能会比最佳状态下下降一个数量级。
53、5.2.3生物发光强度信号的衰减特性:当生物体接受光源刺激,生物发光强度信号迅速到急剧增大并很快达到最大值;然后慢慢衰减,在150s之后光强几乎为零(此过程对应着生物体细胞器能量代谢活性降低出现类似“疲劳现象”以及外部刺激信号的减弱或停止)。在某些实验结果中,经过短暂的时间间隔之后,生物体的受激发发光过程可以被再次启动,但是,生物发光强度信号会弱于前一次生物发光过程。
54、5.3生物体受激发光中单细胞发光数理模型:
55、本文主要探讨受激发的生物体发光过程的生物发光强度信号。基于入射光的生物发光效应,本文建立激光生物学效应的数学物理模型。
56、5.3.1本文所涉及的专业术语如下:
57、生物体发光:生物体在生理或病理机制条件下的发光现象。该过程不仅与外部环境因子相关联,还受制于生物体的内在制约因素(物质或能量代谢状态、基因遗传与免疫应答机制)。发光生物体的发光细胞数量:发光生物体的全部细胞中具有潜在的发光能力的细胞数量:发光生物体的发光能力参数(glowing ability parameter):发光生物体的单个发光细胞所产生的生物光子信号的强度。它集中性地体现了发光生物体的发光生物学特性,也可以解释外部环境因子对于不同生物体发光过程的影响作用呈现出较大的差异性。生物体的发光能力参数是内在机能与时间的函数。对于单基因控制的分子遗传机制,生物体发光(glowing)对应的发光能力参数g(u,t)是两个变量(u与t)的函数,随时间t而改变反映了宏观特性,随内在的机能u而改变反映了生物体的内在制约性机制属性。若细胞的发光能力主要由发光蛋白的生成函数来体现,则发光能力参数g(u,t)可以被简化为发光蛋白的生成函数,其中ga(u,t)与gb(u,t)对应着单一的母细胞或子细胞的发光蛋白的生成函数。
58、激光的发光生物学效应:激光可以作为受激生物发光过程的入射光信号源,通常会产生光效应、热效应、电磁场效应、压力与冲击波效应。本文主要考察其中的可能促进生物发光过程的发光生物学效应。
59、受激发的生物体发光:处于受激发条件下具备了信号加强效应特性的的生物体发光过程。刺激因子可能来自于外部环境因子(例如光信号刺激、电磁信号刺激、动能信号刺激),也可能来自生物体内在信号因子(例如免疫应答信号因子、细胞信号传导因子、细胞信号反馈因子、细胞生长信号因子、基因复制信号因子、基因转录与蛋白翻译信号因子)。外因通过内因起作用,相应地,源自外部环境因子的信号刺激效应受制于发光生物体的内在的生理或病理机制。
60、受激发发光的生物体:在受激发条件下完成生物发光过程的生物体,可以是发光蛋白、发光细胞、发光细菌、发光病毒、发光藻等。
61、受激发的生物体发光过程的内在制约因素:内在制约因素涉及了物质或能量代谢机能状态、基因遗传与免疫应答机制等。
62、细胞发光概率p(glowing probability)指δt时间间隔内单个细胞受激发后产生生物发光强度信号的概率,p=λδt,其中细胞发光能力参数λ(cell glowing abilityparameter)表征了生物学属性中的特异性。
63、细胞的发光功能分化趋势:将现实世界中处于极其微小体积与极其微小时间内的细胞发光功能状态理解为由概率η制约的朝着发光期与间歇期发展的两种功能分化趋势(以概率η分化为具备发光能力的细胞,对应着以概率1-η分化为不发光的细胞)。概率η制约着细胞两种功能分化趋势的大小,并通过概率的差异来集中地反映细胞群复杂的生理或病理作用机制(例如多次发光可能让细胞出现“疲劳”现象、细胞的发光与不发光状态存在双向转化)。
64、发光生物体的不发光概率:发光生物体处于不发光状态下的概率。考虑到发光概率与不发光概率的加和值为1,于是不发光概率=1-发光概率。
65、连续时间段内的不发光概率:等于连续时间段内的各个子时间区域中的不发光概率的乘积。细胞群中发光蛋白的生成函数:如下两个过程制约着发光蛋白的数量:发光蛋白的降解过程(对于被本细胞周期继承的来自上一个细胞周期的发光蛋白)与新生的发光蛋白的形成过程(发生在第n个细胞周期)。时间依赖性边际分布的生成函数g(u,t)是(1+u)nn与p(nn,t)卷积的累积加和值,其中nn从0变化到∞,而p(nn,t)是两个变量(nn与t)的函数。在同一个细胞周期中,ga(u,t)与gb(u,t)分别定义为对应着单一的母细胞或子细胞的发光蛋白的生成函数。对于第n个细胞周期,gn(u,t)={gn(uexp(-kt),0)}*{ga2(u,t)}*{gb2(u,t)},其中gn(uexp(-kt),0)对应着发光蛋白被降解的过程,ga2(u,t)与gb2(u,t)分别对应着母细胞与子细胞中新生的发光蛋白的生成函数。
66、发光生物体的积分边界条件:积分边界条件指初始时刻(t=0)对应着发光生物体的不发光概率为1。在探测时间范围内的初始时刻,发光生物体因尚未受到刺激因而肯定地处于不发光状态。
67、发光能量e(glowing energy)是发光时间与源自外部环境因子的对于发光过程的主要促进因素的函数。
68、发光细胞数量函数θ(q)指某个时刻生物体中具有发光能力的细胞数量,它受细胞周期中细胞生长机制制约因而是随时间变量q发生变化。任何检测设备都设定了背景信号(噪音信号)的临界值,意味着低于此临界值的任何信号都被当成了背景信号。仪器能检测到的生物发光强度信号(在给定的时间范围内)等于发光细胞数量θ在tc时间间隔内的积分。
69、发光信号能量的函数e(t-tc,h(q))的变量因素是时间间隔(t-tc)以及主要刺激因子h(q),其中,(t-tc)扣除了从峰值衰减到临界值的时间tc,而h(q)随时间变量q而变化,并且因发光生物体的生物学特性与受激发发光过程的分子机理具有特异性而呈现显著差异性。生物发光信号的强度i(t):依据微积分定义可以表示为发光细胞数量函数θ(q)与发光能量函数e(t-tc,h(q))的卷积。
70、重积分方式计算生物发光信号的强度:重积分中首先在积分变量q的区间(s-tc,s)积分(从给定的时间长度s中扣除了生物体发光信号从峰值衰减到临界值所需要的时间tc),然后在积分变量s的区间(t-ω,t)积分(全部检测时间内有效地发出生物体发光信号的时间长度是ω)。
71、单细胞受激发生物发光的研究对象:研究单细胞生物发光过程,不仅涉及发光事件的概率性(从统计学意义回答生物体实施生物发光过程“有多大的可能性”,而且涉及了发光事件的执行程度(从时间维度看属于离散性或连续性;从短期效应的数量维度看属于强或弱;从长期效应的数量维度看属于均匀或起伏;从空间维度看属于局部或全面)。
72、单细胞受激发发光的主要促进因素:外部环境因子对于生物体发光过程可能具有促进作用,对应的主要促进因素常常呈现出差异性。光源刺激的情形下“主要促进因素”包括了入射光的频率;电磁刺激的情形下“主要促进因素”包括了电磁场的强度。以光信号刺激为例,主要刺激因素h(q)可以是光的频率h(e=hv),对于电磁信号刺激的情形,主要刺激因素h(q)可以是电磁场的特性参数(电场强度e=f/q,即电场对试探电荷的作用力与试探点电荷带电量的比值;磁感应强度(magnetic flux density)描述磁场强弱和方向,洛伦兹力公式指运动电荷q在垂直于磁场b的方向以速度v运动时所受的磁场力f=qvb)。
73、单细胞受激发发光的累积效应:参照微分定义将时间区域划分成连续的子时间区域,于是整个探测时间段内的生物发光过程看作是多个连续的子发光过程的累积效应。
74、发光细胞群:将局限于δv(体积非常小)的空间范围内的多个细胞看作是属于一个发光细胞群。该细胞群中单个细胞具备了发光能力方面的特异性(体现在细胞发光能力参数λq存在差异)。以单位体积中包含着n0个具有潜在的发光能力的细胞来考虑,δv中累积的发光效应来自n0δv个发光细胞的聚集效应。
75、发光细胞群的序号标注:处于不同细胞阶段的细胞群的序号顺序地采用大于或等于零的整数(即0,1,2,3...)。各序号对应着发光过程的第几次(0指不发光,1指第一次发光,2指第二次发光,依次类推)。这些序号将出现在细胞群各参数的右上角圆括号内(例如λ(0)))。
76、发光细胞群序列间的内在联系:细胞群发光效应综合性地体现了发光细胞群序列中前后序号之间相互作用以及发光细胞与不发光细胞之间的相互转化。从数量角度看,以相邻整数作为序号标注的每个细胞群中的细胞数量受制于相邻的细胞群以及细胞群的数量转换机制的特性。从能量角度看,以相邻整数作为序号标注的每个细胞群中的细胞发光能量受制于相邻的细胞群以及细胞群的能量转换机制的特性。
77、发光细胞群的细胞数量边界条件:微分方程组中关于细胞数量的边界条件指在初始时刻(t=0)第m号细胞群中包含的细胞数量n(m)(0)=n0(m),其中表达式中字母n右上方指数部分的字母m对应着第m号细胞群,等号右方字母n右下角的下标0指初始时刻(t=0)。
78、基因失活率与基因激活率:σb基因失活率与σu基因激活率分别对应着细胞处于不响应时期与生物发光时期。
79、关于基因长时间失活的假定:本文将复杂的基因密码遗传过程简化地描述为基因激活与基因失活两种状态之间转换。基于细胞是大概率地处于不响应时期的统计数据,本文假定细胞周期中基因长时间处于失活状态。实验结果表明绝大多数生物体发光过程中大部分时间都处于off状态(δ=σb/σu即基因失活率除以基因激活率,其比值较大)。基因失活利于避免基因复制与基因转录发生冲突,它是漫长的物种进化过程向幸存的物种赋予的一种利于机能保护性与机能促进性的生理机制。
80、二项分布(binomial distribution):即随机变量x的离散概率分布。在n次独立重复的伯努利试验中设每次试验中事件a发生的概率为p,用x表示n重伯努利试验中事件a发生的次数,则x的可能取值为0,1,…,n,且对每一个k(0≤k≤n),事件{x=k}即为“n次试验中事件a恰好发生k次”。
81、细胞周期:包含分裂间期(interphase)和分裂期(mitotic phase或divisionphase)。细胞分裂(cell division)指活细胞增殖及其数量由一个细胞分裂为两个细胞。分裂前的细胞称母细胞(mother cell),分裂后形成的新细胞称子细胞(daughter cell)。
82、5.3.2单细胞受激发生物体发光的微分方程数学模型:
83、5.3.2.1单细胞的生物发光强度信号的累积效应:
84、如果把时间段t分割成m个连续的子时间段(m是大于零的整数,m=t/δt),于是整个时间段内的生物发光过程可以看作是m个连续的发光过程的累积效应。则时间段t内不发光的概率是∏(1-λiδt)=(1-λ1δt)*(1-λ2δt)*(1-λδt)......(1-λnδt),当δt很小,上式只需要计入一阶导数,(1-λiδt)=exp(-λiδt),于是p=exp[(-λ1δt)+(-λ2δt)+(-λ3δt)+...+(-λnδt)]=exp[δt(-λ1)+(-λ2)+(-λ3)+...+(-λn)]=-exp[δt(λ1+λ2+λ3+...+λn)],依据定积分的定义,被积分函数λq的定积分对应着全部的λqdq在区间内的加和。将λqdq中dq换成均等的δt,则变成了∫(0,t)(λq)dq。因此δtσ(λ1+λ2+λ3+...+λn)=δt(λ1+λ2+λ3+...+λn)=∫(0,t)(λq)dq,当δt趋近于零,p=exp[-∫(0,t)(λq)dq]。
85、关于发光生物体的积分边界条件:从上式可知,当t=0,发光生物体的不发光概率为1(在初始时刻生物体因尚未受到刺激因而肯定地不会发光)。
86、关于受激发生物发光过程的内在制约因素:当t>0,生物体的不发光概率呈现依据指数规律下降的趋势,下降的特性主要受制于细胞发光能力参数λ(cell glowing abilityparameter)(细胞发光能力参数λ作为变量系数出现在指数部分)。从上边的数学模型可以得知,生物体发光这个过程对应的发光概率受制于细胞内在的特性(细胞发光能力参数λ),表明了生物体发光的内在制约因素(物质或能量代谢机能状态、基因遗传与免疫应答机制)是生物发光强度信号的关键性制约因素。
87、5.3.2.2单细胞的生物发光强度信号的背景信号效应:
88、本文引入了发光细胞数量函数θ(q)以及发光信号能量函数e(t-tc,h(q))。
89、发光细胞数量函数θ(q)指某个时刻生物体中具有发光能力的细胞数量,它受细胞周期中细胞生长机制制约因而是随时间变量q发生变化。将生物发光强度信号从峰值衰减到临界值ic所需要的时间定义为tc,仪器能检测到的生物发光强度信号(在给定的时间范围内)都必定是高于背景信号(噪音信号)的临界值ic。
90、发光信号能量的函数e(t-tc,h(q))的变量因素是时间间隔(t-tc)以及主要刺激因子h(q),其中,(t-tc)扣除了从峰值衰减到临界值的时间tc,而h(q)随时间变量q而变化,并且因发光生物体的生物学特性与受激发发光过程的分子机理具有特异性而呈现显著差异性。将能发光的细胞与不发光的细胞进行了区分,同时将采用仪器检测到的生物发光强度信号扣除了低于临界值的部分n(t)从而留下了超出临界值的部分。
91、生物发光信号的强度i(t)可以依据微积分定义表示为发光细胞数量函数θ(q)与发光能量函数e(t-tc,h(q))的卷积,i(t)=∫(t-tc,t)θ(q)e(t-tc,h(q))dq此处将积分区间的起始时间设为t-tc利于扣除背景信号,意味着给定的时间范围内包含着一个信号从峰值衰减到临界值的时间tc。卷积θ(q)e(t-tc,h(q)中发光细胞数量θ(q)是积分变量q的函数,发光能量函数e(t-tc,h(q))是时间维度变量(t-tc)以及主要刺激因子h(q)的函数。主要刺激因子h(q)是积分变量q的函数,是促进单个细胞的生物发光强度信号的因素。
92、把背景信号为零作为边界条件:一般情况下由θ=n(t-tc)-n(t)可知,n(t-tc)是(t-tc)时刻对应的生物发光强度信号效应,需要减除其中包含的低于临界值ic的生物发光强度信号。上边θ=n(t-tc)-n(t)考虑了潜在的具有发光能力的细胞数量,以及合适的时间范围(积分区间位于t-tc与t之间)。当不考虑背景信号(噪音信号)的干扰性时,所有的发光强度信号都可以被仪器检测并被保存为有效信号;将生物发光强度信号从峰值衰减到临界值ic(把ic定义为0)所需要的时间定义为tc(等于t)。意味着,在给定的时间范围内,生物发光强度信号尚未从峰值衰减到临界值ic,一直到给定的时间范围的最后一刻,生物发光强度信号刚好衰减到临界值。θ=n(t-tc)-n(t),当tc等于t时n(t-tc)对应着n(0)。于是θ=n(t-tc)-n(t)=n(0)-n(t)=0-n(t)=-n(t)。仪器检测的结果是即时效应,n(t)对应着t时刻的生物发光强度信号效应)。(t-tc)时刻对应的信号强度不同于(t)时刻的信号。
93、5.3.2.3单细胞的生物体发光信号的强度:
94、下边采用两变量的重积分方式计算生物发光信号的强度:
95、关于单细胞不存在多次发光现象的假定:θ(q)是λ(q)与n(q)两者的函数。本文在下边的简化情形中认为发光细胞不存在多次发光现象,并且在整个探测时间内细胞参数λ(q)与n(q)都仅仅是时间变量q的函数,于是细胞发光能力参数λ(q)与细胞数量n(q)的乘积就等于发光生物体的有效发光细胞数量。
96、首先在积分变量q的区间(s-tc,s)积分,然后对于积分变量s的区间(t-ω,t)进行积分。重积分中首先在积分变量q的区间(s-tc,s)积分意味着对于(s-tc,s)积分中扣除了背景信号(噪音信号),原因是从给定的时间长度s中扣除了发光强度信号从峰值衰减到临界值所需要的时间tc(在生物发光时期中包括了信号高于临界值与低于临界值两种情形)。
97、重积分中在积分变量s的区间(t-ω,t)积分意味着整个探测时间的时间长度t中包含了不响应时期以及生物发光时期,全部检测时间内对于生物发光信号发挥作用的时间区域是(t-ω,t),对应的时间长度是ω。
98、i(t)=∫(t-tc,t)θ(q)e(t-tc,h(q))dq表明了生物发光强度信号的强度i(t)对应的被积分函数(发光细胞数与发光能量函数的卷积)是时间t的函数,积分结果即强度i(t)也是时间t的函数。
99、i(t,ω)=∫(t-ω,t)∫(s-tc,s)θ(q)e(t-tc,h(q))dqds重积分中首先在积分变量q的区间(s-tc,s)积分,其中s指时间变量,s-tc指扣除了发光强度信号从峰值衰减到临界值所需要的时间tc;重积分中然后在积分变量s的区间(t-ω,t)积分,其中积分时间区间的时间长度ω指探测器在单位体积中检测到发光强度信号所耗费的时间,t是整个探测时间的时间长度。
100、应用θ(q)=λ(q)n(q),生物发光强度信号i(t,ω)=∫(t-ω,t)∫(s-tc,s)λ(q)n(q)e(t-tc,h(q))dqds。概括以上分析,具有普遍意义的表达式表明仪器检测结果显示的生物发光强度信号i(t,ω)对应着积分时间区间的时间长度ω以及整个探测时间的时间长度t。
101、5.3.3细胞群的生物体发光信号的强度
102、细胞数量越多,累积的发光效应越强。为简化论述,将局限于δv(体积非常小)的空间范围内的多个细胞看作是属于一个细胞群。该细胞群中单个细胞具备了发光能力方面的特异性(体现在细胞发光能力参数λq存在差异)。以单位体积中包含着n0个具有潜在的发光能力的细胞来考虑,δv中累积的发光效应来自n0δv个发光细胞的聚集效应。考虑生物发光过程中细胞群体的累积性发光效应可以分为时间维度上的短期效应与长期效应。对于某一个确定时间点的短期效应,将细胞群体中单个细胞的发光效应累积起来就得到整体效果。
103、发光细胞群的发光概率:参照微积分的通常假定,在体积非常小的空间范围内把细胞的体积密度定义为n0。为便于采用最简单的方式计算细胞群的发光概率,假定该细胞群中所有细胞都拥有相同的发光概率,因而全体细胞中能发光的细胞数量等于细胞数乘上对应的发光概率。n=n0p=n0exp[-∫(0,t)(λq)dq]上边表示了细胞群的整体发光特性是单个发光细胞的细胞参数λq在时间t内的积分效果。
104、关于细胞群中单个细胞最多发光一次的假定:此处假定了在仪器探测的时间范围内“发光细胞不存在多次发光现象”,意味着潜在的具有生物发光能力的细胞完成一次生物发光过程之后即进入不响应时期(不再具有发光能力)。这个假定仅仅为了便于简化关于发光概率的计算(潜在的具有发光能力的细胞拥有确定的发光概率)。在所研究的时间范围内假定所有细胞最多有一次发光机会(发光过程后进入到绝对不响应时期)。发光细胞在执行发光任务之后被排除在发光能力范围之外了(发光概率不会多次地或重复地产生作用);细胞群受到外界刺激后的累积发光效应呈现出指数衰减规律(细胞发光能力参数λ作为时间t的系数出现在指数部分)。
105、对于生物发光过程的短期效应,在时间范围非常小的情况下,可以认为单个细胞的细胞发光能力参数λq不随时间发生改变。由n=n0p=n0exp[-λt]可知,细胞群中由发光概率对应着潜在的具有发光能力的细胞数量;
106、为考察边界条件,初始时刻t0,单位时间单位体积内的发光细胞数量θt0=-dn/dt(t=t0)=(λt0)n0exp[-∫(0,t0)(λq)dq]=(λt0)nt0θt0指初始时刻t0对应的单位时间单位体积内的发光细胞的数量。给定了初始时刻t0,单位时间单位体积内的发光细胞数量与下述变量成正比关系(单个细胞的细胞参数λt0以及潜在的具有发光能力的细胞数量nt0)。
107、5.3.4关于细胞群中单个细胞实施多次发光过程的分析:
108、在现实世界中,在仪器探测的时间范围内发光细胞确实可能存在多次发光现象;不同的发光细胞处于细胞周期的不同阶段(发光期或间歇期),并不会步调一致地处于相同的阶段。
109、为了便于标注处于不同细胞阶段的细胞群,顺序地,细胞群的序号采用大于或等于零的整数,即0,1,2,3,(对应的各个序号指处于发光过程的第几次(0指不发光,1指第一次发光,2指第二次发光)。这些序号将出现在下边各个参数的右上角的圆括号内(例如λ(0))。
110、以发光序号标记细胞群:通过对多次发光过程采用顺序地标注第几次发光,细胞发光能力参数λ、细胞数量n与发光概率等特性被赋予给相应的细胞群。基于多次发光过程中后边的发光细胞都经历了早先的细胞过程(早先处于发光状态或不发光状态),本文顺序地指定细胞群的序号(对应着发光顺序的先后)。过程的先后顺序制约了细胞群的变化发展方向(细胞的数量随着过程进展而依据概率大小下降)。细胞发光能力参数λ(反映细胞内在特性)随着过程进展而出现某种程度的下降(实验结果表明了多次发光将因消耗细胞物质与能量而呈现出类似“疲劳”的机能耗损现象)。有益效果是,对于不同过程之间的衔接(包括发光过程与发光过程之间,以及发光过程与不发光过程之间),细胞数量的静态计算(例如λ(1)n(1))依据了细胞发光能力参数(λ)与细胞数量(n);细胞功能分化的数量计算(例如η(1)λ(1)n(1))考虑了概率大小(例如在全部细胞中依据概率η(1)获得具有生物发光功能的细胞数量)。在单位时间单位体积内如下数量(λ(0)n(0))的细胞将从第0号细胞群(处于不发光的间歇期)进入到第1号细胞群(将实施第1次发光);若将这个转变过程对应的发光概率记为η(0),依据发光概率可知其中真正有潜在的发光能力的细胞数量为η(0)λ(0)n(0),而(1-η(0))λ(0)n(0)表示了初次发光的细胞中进入不响应时期的细胞数量。
111、在单位时间单位体积内如下数量(λ(1)n(1))的细胞将从第1号细胞群(对应第1次发光)脱离出来;若将这个转变过程对应的发光概率记为η(1),依据发光概率可知其中真正有潜在的发光能力的细胞数量为η(1)λ(1)n(1),而(1-η(1))λ(1)n(1)表示了从第1号细胞群(完成第1次发光)进入不响应时期的细胞数量。
112、细胞发光功能分化趋势:在具有普遍意义的细胞生长或代谢进程中,在指定的极其微小的时间维度与空间维度上,发光与不发光这两种状态可以相互转化(转化的可能性对应着概率η或概率1-η)。将现实世界中处于极其微小体积与极其微小时间内的细胞功能状态(发光期与间歇期)理解为由概率η制约的两种功能分化趋势(以概率η分化为具备发光能力的细胞,对应着以概率1-η分化为不发光的细胞)。概率η制约着细胞两种功能分化趋势,并通过概率的差异来集中地反映复杂的多种作用机制(例如多次发光可能让细胞出现“疲劳”现象、细胞的发光与不发光状态存在双向转化)。
113、前后发光过程之间的衔接:细胞发光现象涉及了前后发光过程之间的衔接以及发光细胞与不发光细胞之间的相互转化。以相邻整数作为序号标注的每个细胞群中的细胞数量受制于相邻的细胞群以及衔接机制的特性。
114、递推表达式为dn(m)/dt=η(m-1)λ(m-1)n(m-1)-λ(m-2)n(m-2)具体的书写形式是,对于第0号细胞群,dn(0)/dt=-λ(1)n(1)这里的符号是表征数目是呈现下降趋势;对于第1细胞群,dn(1)/dt=η(0)λ(0)n(0)-λ(1)n(1)对于第二号细胞群,dn(2)/dt=η(1)λ(1)n(1)-λ(2)n(2)。
115、发光细胞群的细胞数量边界条件:上述微分方程对应的边界条件如下:在初始时刻t=0第m号细胞群中包含的细胞数量n(m)(0)=n0(m),其中表达式中字母n右上方指数部分的字母对应着第m号细胞群,字母n右下角的下标0指初始时刻t=0对应的细胞数量;具体的书写形式是,在初始时刻t=0第0号细胞群中包含的细胞数量n(0)(0)=n0(0),在初始时刻t=0第1号细胞群中包含的细胞数量n(1)(0)=n0(1)
116、计算机软件系统可以通过逐级迭代法求解上述微分方程组。通常的求解过程包含了对于单个微分方程进行变形(例如,将对应于第m号细胞群的方程两边同时乘上积分因子∫(0,t)(λ(m)(p)dp,并在后续的积分过程中应用边界条件n(m)(0)=n0(m))。细胞发光能力参数λ是制约生物体发光特性的内在属性(对于随时间变量p发生变化的λ采用λ(m)(p)来表示,其中上标m表明细胞属于第m号细胞群)。
117、5.4关于细胞发光能力参数的数学模型:
118、上述数学模型中将多种复杂的生物机制的内在制约性作用集中地由细胞发光能力参数λ来体现。考虑到生物体的生物发光现象对应着无数复杂的生物机制,有必要深入探讨关于细胞发光能力参数λ的求解过程。细胞周期中存在分裂与复制的过程;过程中涉及基因密码遗传与代谢调控等分子机理(例如,基因遗传密码中启动子发生转换、生长依赖性转录、单基因拷贝完成后激发新生的发光蛋白)。新生的发光蛋白的数量是关于时间变量的函数,对应的涉及时间依赖性边际分布的生成函数g(u,t)是(1+u)nn与p(nn,t)卷积的累积加和值,其中nn从0变化到∞。p(nn,t)是两个变量(nn与t)的函数。
119、5.4.1生物体的发光能力参数是内在机能与时间的函数。对于单基因控制的分子遗传机制,生物体发光(glowing)对应的发光能力参数g(u,t)(即上文所述的细胞参数λ)是两个变量(u与t)的函数,随时间t而改变是宏观层面的现象,随内在的机能u而改变反映了生物体的本身机制属性。发光蛋白分子的初始边际分布p(i)=pi意味着g(u)=∑ipi(u+1)i。考察启动子转化与新生的发光蛋白(时间依赖性的基因转录几率)等因素,受制于细胞生长状况的基因转录过程对应的随机动力学可以采用如下方式进行描述:在单步骤反应中,新生的发光蛋白的生成速率为σu(基因激活率),依据二项式分布,其平均的时间依赖性尺寸分布遵循(ρ0/σb)exp(αt)。
120、5.4.2关于细胞周期中基因长时间处于失活状态的假定:本文将复杂的基因密码遗传过程简化地描述为基因激活与基因失活两种状态之间转换。在基因复制过程的开始时刻,细胞到底是对应着基因激活与基因失活两种状态之中的哪一种状态,不得而知。统计数据中大概率事件是细胞处于不响应时期。σb基因失活率与σu基因激活率分别对应着细胞处于不响应时期与生物发光时期。实验结果表明,绝大多数生物体的发光过程的特征是大部分时间都处于off状态(δ=σb/σu即基因失活率除以基因激活率,其比值较大)。off状态时间比较长是漫长的物种进化过程向幸存的物种赋予的一种利于机能保护性与机能促进性的生理机制。off状态时间比较长对于维护细胞的正常生理机制具有非常重要的意义,为生物体的发光过程进行物质储备以及能量储备等一系列准备工作。这类似于细胞周期中分裂间期时间比较长而分裂期比较短。复制过程的结果是形成了母细胞的基因组以及子细胞的基因组。子细胞可以是延续母细胞的处于基因激活状态,也可以是转入到基因失活状态。对于具有紧密关联作用的母细胞及其子细胞,实验研究结果表明了在基因复制过程中以及复制过程刚好结束时基因转录过程并没有发生。客观上,细胞转入到基因失活状态具有了重要的生物学意义,可以避免基因复制过程与基因转录过程发生冲突(利于维持dna结构的稳定性并防范基因密码发生错失)。在子细胞的复制前期不存在基因转录过程,因此[0,tr)时间范围内gb(u,t)=1。
121、5.4.3发光蛋白的生成函数:把ga(u,t)与gb(u,t)分别定义为对应着单一的母细胞或子细胞的发光蛋白的生成函数(处于一个细胞周期中)。td指细胞分裂间期(cellularinterdivision time)。tr是复制所耗费的时间(因为复制所耗费的时间远远小于细胞周期对应的时间长度,可以近似地认为细胞复制过程是瞬间完成的)。在时间区间[0,tr)内,ga(u,t)对应着处于细胞周期的基因复制前期状态下采用随机动力学描述的新生的发光蛋白的生成速率。在时间区间[tr,td)内,ga(u,t)对应着处于细胞周期的基因复制后期状态下采用随机动力学描述的新生的发光蛋白的生成速率。ρ1=ρ0exp(αt)指基因复制时转录速率;ρ0指细胞周期起始时刻的转录速率;α=(1/td)ln(vf/v0),其中v0是细胞周期初始时刻的细胞体积,vf是细胞即将开始分裂前的细胞体积。当转录过程不随着细胞生长过程而发生改变,ρu(t)只是时间变量的函数,因α=0因而ρu(t)=ρ0exp(αt)=ρ0。
122、基于实验结果,假定细胞体积随着细胞年龄时间t呈现指数型增长(细胞年龄时间t处于0与细胞分裂间期td这两个数值之间)。假定成熟的发光蛋白按照一阶动力学规律以速率d被降解(关于一阶动力学特点的假定与实验结果是基本吻合的)。不同时期(来自于基因复制前期或基因复制后期)对应的新生的发光蛋白的生成速率具有独立性特点。
123、5.4.4相邻细胞周期中发光蛋白的生成函数:p(nn,t)指新生的发光蛋白在第n个细胞周期中时刻t的边际分布。考察两个生成函数(在第n个细胞周期的初始状态下的生成函数,在第(n-1)个细胞周期的时刻td的生成函数),对于给定的第n个细胞周期中某具体的细胞年龄时间t,有如下两个过程制约着发光蛋白的数量:发光蛋白的降解过程(对于被本细胞周期继承的来自上一个细胞周期的发光蛋白)与新生的发光蛋白的形成过程(发生在第n个细胞周期)。gn(u,t)=gn(uexp(-kt),0)ga2(u,t)gb2(u,t),gn(uexp(-kt),0)对应着发光蛋白被降解的过程,ga2(u,t)对应着母细胞中新生的发光蛋白的生成函数,gb2(u,t)对应着子细胞中新生的发光蛋白的生成函数。gn(u,0)=gn-1(ηu,0)ga2(u/2,td)gb2(u/2,td)n>1;η=[exp(-ktd)]/2。在求解动力学过程之前需要赋予一个确定的数值给g1(u,0)(指第一个细胞周期的初始状态)。考虑到初始状态下新生的发光蛋白的生成速率为零,本文指定g1(u,0)=1。实验研究结果充分地表明了新生的发光蛋白在瞬间转化为成熟的发光蛋白k>>d(实测数据举例,k=1.25min-1d=0.0462min-1,k/d=27.06);基因在绝大多数时候处于基因失活状态(基因失活数量与基因激活数量之比δ>>1)。新生的发光蛋白在瞬间转化为成熟的发光蛋白,意味着新生的发光蛋白快速地拥有了生物发光功能并进入到生物发光状态。这里表明两个阶段(新生的发光蛋白与成熟的发光蛋白)之间的时间间隔非常短,但是这两个阶段彼此独立存在并遵循各自的动力学规律。
124、5.4.5对于成熟的发光蛋白发生波动的相关系数的敏感性分析如下:可以考察sd除以平均值。处于细胞年龄时间t的细胞群拥有确定数量的发光蛋白分子,这与各个发光蛋白分子分别属于第几个细胞周期是没有关联性的。gn+1(u,t)=gn(u,t)考虑在整个细胞周期内的平均值,成熟的发光蛋白在处于细胞稳态条件下的相关系数cv=td-1∫(0,td)sqrt{[δ2(nm,t)]/(nm)t}dt,数值计算方法是将一个细胞周期的时间均等地分割为100份。cv对于某个参数r的相关敏感性可以表示为意味着某个参数r产生1%的数值变动将引起cv出现λ%的数值波动。胞信使rna以速率被翻译成发光蛋白;发光蛋白以速率dp被降解;此两过程都遵循一阶动力学规律。实验研究结果表明,信使rna的被降解速率的平均值大约是发光蛋白被降解速率的三倍到五倍(例如,基于超过四千个基因的实证研究结果,对小鼠为三倍,对酵母为五倍)。因此,对于数千个基因,可以假定k>>d>>dp,意味着基因过程比蛋白过程更快捷迅速更有效率(与发光蛋白关联的新生基因与成熟基因对应着生成函数,所耗费时间远小于新生的发光蛋白与成熟的发光蛋白对应的时间)。实验研究结果表明:对于以实验动物小鼠的胚胎干细胞中相当数量(例如超过一千)的基因作为测试对象,都具有基因失活时间区间为主的特性(基因失活数量与基因激活数量之比δ>>1,例如δ取值大于5)。cv对于某个参数r的相关敏感性研究结果显示,最敏感的参数是发光蛋白的降解速率d与剂量补偿性参数σu(b)与σu(a)(σu(b)对应着复制之前,而σu(a)对应着复制之后);而敏感性最低的参数是对细胞生长过程具有依从性的转录参数以及复制时间tr
125、5.5关于发光生物体生命周期的质量控制体系
126、考虑到发光生物体的生命周期中遗传代谢机制存在许多不确定因素,现实世界中难以准确判断是否满足细胞发光的特性指标(“细胞分裂间期td”、“复制所耗费的时间tr”、“发光能力参数”、“基因失活数量与基因激活数量之比δ”、“母细胞中新生的发光蛋白的生成函数”、“子细胞中新生的发光蛋白的生成函数”、“发光细胞群的细胞数量边界条件”)。本发明通过质量控制体系追踪并求解受激生物发光过程的动力学参数从而实现“智能化控制受激生物发光过程”。
127、本发明采用数学模型阐释了细胞受激发光过程的动力学条件,有益效果是,利于通过参数指标来区分细胞受激发光过程的动力学阶段。优选地,通过求解“发光能力参数”、“发光蛋白的生成函数”等参数指标可以优化细胞受激发光过程的动力学特性从而打开或关闭细胞遗传代谢机制的信号通路,将利于以细胞受激发光过程的特性光谱信号优化激光制导导弹技术的信号编码调制。数学模型以细胞群之间的关联作用反映“生物光子链式反应机制”,依据属于多次发光过程中第几次发光而顺序地标注细胞群,并考察相应的细胞发光能力参数、细胞数量与发光概率等特性。
128、5.6受激发生物光子信号对于激光制导导弹技术的重要意义
129、5.6.1提升激光信号的编码加载与激光信号的解调识别能力
130、激光制导导弹系统中激光信号接收器所探测的回波信号可能受到多种因素的干扰。开发适用于激光制导导弹系统的具有“信号保真性能高、抗信号干扰能力强”特点的源自生物体受激发光过程的信号源,利于提升激光制导导弹系统中激光信号的编码加载与激光信号的解调识别能力,从而利于提升军事侦察的灵敏度与准确度并大大提高精准打击能力。
131、生物光子信号发射器利于提升信号识别能力:基于生物体发光细胞的遗传代谢机制中“关闭”与“激活”两种工作状态可以相互转换的特点,军事侦查目标对应的多种连续性信号能够被转换为数字信号,于是大大增强了激光制导导弹系统的信号编码调制与信号解调识别能力。
132、5.6.2打开或关闭细胞遗传代谢机制
133、本发明的特点是以理论推导的结果(细胞受激发光过程的动力学特性)来促进生物光子信号的产生过程以打开或关闭细胞遗传代谢机制的信号通路,利于筛选发具有优秀发光能力的发光生物体并改善生物光子信号的光谱特性,从而利于优化激光制导导弹技术的信号编码调制。本发明的技术策略是通过评判宏观层面的综合性性能优劣来调控微观层面的运行机制,这是以结果为导向来调控内在运转机制。生物医学领域中的疾病防治策略采取以综合性机能指标为导向来优化药物或手术治疗方案。从军事工业科技进步的角度看,本发明指出了生物体受激发光技术与激光制导导弹系统的融合发展,将通过示范性作用与产业化升级过程带动一系列生物医学高端技术融入到国防科技产业领域。
134、5.6.3易于编码调制的阶跃性数字信号
135、现代军事战争对于军事侦查目标的信号准确度与精密度提出了越来越高的技术要求,常规的信号编码调制与信号解调识别技术具有很大的发展空间。借助计算机网络系统的数字信号传输过程,本发明将生物发光强度信号实施“信号加载”与信号传输”后具备了“高度保真”、“高效传输”、“高效破译”等特性。
136、通常,细胞的生理病理过程接收外部的环境因子给予的信号刺激属于模拟信号,细胞通过信号反馈机制所产生的生物发光强度信号也是模拟信号。生物体发光细胞可以被看作是精巧的高精度信号转换装置,可以将连续性信号转换为许许多多阶跃性的数字信号。输入不同能量层级的信号必定对应着“能够激活细胞发光”与“不能激活细胞发光”两种刺激效应中的一种。从实施角度看,通过采用计算机软件系统的二进制代码,本发明提高了数据传输与数据编码过程的效率。基于“基因遗传密码”与“细胞代谢调控”等生理病理机制,本发明借助细胞的高效识别能力,将复杂的军事信息由模拟信号转变为数字信号。军事侦查目标对应的多种连续性信号需要借助计算机软件系统被区分成许多不同能量层级的信号,然后采用人工智能神经网络系统将能量层级差异性很小的连续性信号用于照射合适的发光细胞,借助细胞信号反馈过程中“能够激活细胞发光”与“不能激活细胞发光”两种刺激效应的差异性把连续性信号区分为阶跃性信号。将生物体发光细胞的受激发光过程融入信号编码调制与信号解调识别技术,将突破性地把连续性信号转化为阶跃性的数字信号,利于增强激光制导导弹系统的信号编码调制与信号解调识别能力。本发明的实施方案阐释了将模拟信号转变为数字信号,其有益效果是,不仅拓宽了激光制导导弹系统中复杂军事信息的采集范围,而且提高了复杂军事信息的“信号加载”与“信号传输”能力。
137、5.6.4“生物光子信号发射器”的关键技术环节
138、“生物光子信号发射器”的关键技术环节涵盖了“发光细胞的筛选”、“细胞发光能力的优化”、“遗传代谢机制中关闭与激活两种工作状态的转换”。本发明提出四个实施步骤:“信号区分步骤”指连续性信号需要借助计算机软件系统被区分成许多不同能量层级的信号;“信号刺激步骤”采用人工智能神经网络系统将能量层级差异性很小的连续性信号用于照射合适的发光细胞;“信号转换步骤”借助细胞信号反馈过程中“能够激活细胞发光”与“不能激活细胞发光”两种刺激效应的差异性把连续性信号区分为阶跃性信号;“编码调制与解调识别步骤”指将生物光子信号的光谱特性融入到激光制导导弹系统的信号编码识别过程。
139、本发明证实了“生物光子信号提升了激光制导导弹系统的信号传输与信号编码过程中高保密性与高保真性”。特定细胞可以被看作是“高效能的信号接收装置”与“高效率的信号转换装置”,整个过程的内在规律性是细胞生物学与分子遗传学领域的“基因遗传密码”与“细胞代谢调控”等生理病理机制。具体而言,具有复杂生命特征的细胞对于外部的环境因子具有灵敏的感知能力与信号反馈能力。宏观世界(比如潜在的敌方军事力量开展的军事侦查甚至军事入侵活动)存在着信号产生与信号传输过程,相应的的外部军事信息可以被收集,然后通过信号刺激激发特定细胞的某种生物发光潜能。于是,在经历了细胞的信号感知与信号反馈过程之后,外部的军事信息由模拟信号转变为数字信号。
140、6一种调控光敏蛋白并编码受激发生物光子信号的激光制导导弹,其特征之六在于,受激发生物光子可以为激光制导导弹提供基于激光谐振机制的生物体激光发射方法。本文介绍了生物光子智能发射器的技术要素(纳米尺度的激光谐振腔)并采用实例证实了生物光子智能发射器的可行性。关于发光生物体激光谐振腔运行机制的数学模型阐释了激光谐振腔的稳定性条件(“信号正反馈条件”以及“增益系数超出临界值”)。
141、6.1下边列出了相关的专业术语:
142、激光振荡条件(laser oscillation condition):激光器的振荡阈值(oscillationthreshold)指产生激光振荡的最低限制条件。谐振腔的损耗和激活介质的增益决定了激光器是否能够产生自激振荡。
143、自激振荡:产生自激振荡必须同时满足两个条件:1、幅度平衡条件指|af|=1;2、相位平衡条件指其中,a指基本放大电路的增益(开环增益),f指反馈网络的反馈系数。起振必须满足|af|略大于1的起振条件。
144、信号正反馈:反馈信号的极性与系统输入信号的极性相同,通过信号叠加效应增强了系统净输入信号。
145、增益系数:增益系数g描述光强经过单位距离后的增长率。当激光工作物质处在粒子反转状态时,一束光强为i单色光入射后,由于受激辐射作用,光强会不断被放大。
146、粒子数反转(population inversion)指处在激发状态的成员数量比起处于较低能级状态的成员更多。
147、光泵(optical pump)指使激光工作物质发生粒子数反转并产生受激辐射的激励光源。
148、平面激光诱导荧光plif(planar laser induced fluorescence):基本原理是待测环境(如火焰)中痕量组分的分子或原子在入射激光的作用下由低能态跃迁到高能态,能级差与入射光波长共振,以达到在高能态布聚的理想效率,为获得高强度的荧光信号提供前提,随后部分粒子从受激的高能态回落到不同的低能态,从而辐射出由不同波长组成的荧光光谱带,通过总荧光强度测量实现流场可视化。
149、矩阵的分拆表示法:a(:,j)表示取a矩阵的第j列全部元素;a(i,:)表示a矩阵第i行的全部元素;a(i,j)表示取a矩阵第i行、第j列的元素。
150、6.2生物光子智能发射器是采用发光生物体制备的激光谐振腔。
151、生物光子智能发射器的技术要素:把水母细胞中的绿色荧光蛋白引入到生物体细胞中,将激光谐振腔植入单细胞内,然后用谐振腔增强发光。激光谐振腔是激光器的重要组成部分(主要功能有正反馈、谐振、容纳工作物质和输出激光的作用)。激光谐振腔在细胞内形成一个微小的泡(当一束激光照射时激光谐振腔受照射后产生谐振,谐振腔内的荧光物质就会发光。发出的光的波长取决于泡的尺寸大小与折射率。当满足能量补偿条件与稳定驻波条件,来回振荡与受激辐射的连锁反应可以使激光谐振腔产生稳定驻波的激光。受激辐射与吸收过程及自发辐射存在竞争关系。处于激发态的能级原子可以通过自发辐射和受激辐射回到基态,而能量补偿利于实现在光传播的方向上受激辐射不断得到放大和加强。当受激辐射过程超出吸收过程,受激辐射产生振荡并实现粒子数反转。激光谐振腔涵盖处于轴线上的两块反射率很高的镜(全发射镜将光几乎全反射,部分反射镜能发射大部分光而透射少量光)。沿轴向的光子在谐振腔内由于两端反射镜的反射不至逸出谐振腔,被反射回到增益介质的光沿轴线方向继续诱发新的受激辐射,从而使轴向行进的光子不断得到放大和振荡。受激辐射过程中发射出的光子与引起受激辐射的光子具有如下共性特点:相同的频率、发射方向、偏振状态和相位。结果是谐振腔内沿轴向的光强急剧增加并从部分反射镜一端输出。
152、6.3采用发光生物体制备激光谐振腔的步骤
153、6.3.1制备纳米尺度的激光谐振腔
154、作为例子,本文采用聚苯乙烯材料构建纳米尺度的激光谐振腔。聚苯乙烯滴含有荧光染料。内置的荧光染料包括但不限于四甲基异硫氰酸罗丹明,异硫氰酸荧光素(fluoresceinisothiocyanate,fitc)、四乙基罗丹明(rhodamine,rib200)。内置的荧光染料将具有确定的质量浓度或体积浓度,它在激光信号刺激下发出荧光信号,其信号强度在针对荧光信号的定量分析过程中被用作参照性荧光信号)。
155、合成纳米聚苯乙烯:
156、关于聚苯乙烯的通用的合成步骤如下:先将聚乙烯吡咯烷酮溶于苯乙烯与乙醇的混合体系中,然后加入偶氮二异丁腈与表面活性剂。在惰性气体保护下加热将制得乳白色聚苯乙烯(过程中滴加甲基丙烯酸与乙醇的混合溶液)。
157、关于聚苯乙烯的结构修饰存在差异化的操作步骤。作为一个非限制性的例子,用于修饰聚苯乙烯微球表面的操作步骤如下:在搅拌条件下以聚乙烯亚胺与乙醇分散上述聚苯乙烯,添加功能性添加剂(例如多羧基有机酸中的柠檬酸、d-苹果酸、l-苹果酸或dl-苹果酸)以获得表面连有羧基的聚苯乙烯固体微球。若对此聚苯乙烯固体微球作进一步结构修饰(例如制备含有稀土元素的纳米粒子),可以添加稀土盐水溶液(或碱金属盐水溶液)并通过加热搅拌使固体微球重新分散于水中。作为参考,聚乙烯吡咯烷酮分子量可选为55000。加热反应温度为55-75℃,反应时间24-36h。稀土盐可选硝酸铽、硝酸铕、氯化铽、氯化铕;碱金属盐可选磷酸二氢钠、磷酸二氢钾、氟化钠、氟化钾、磷酸钠、磷酸钾。
158、6.3.2植入纳米尺度的激光谐振腔
159、将纳米尺度的激光谐振腔植入发光生物体的单细胞内。激光谐振腔可以在细胞内形成一个微小的泡。作用机制是:当一束激光照射时激光谐振腔受照射后产生谐振,谐振腔内的荧光物质就会发光。发出的光的波长取决于泡的尺寸大小与折射率。
160、本文以聚苯乙烯滴作为纳米尺度的激光谐振腔的例子。将聚苯乙烯滴注射到细胞内。调试过程中可改变聚苯乙烯滴的特性参数(例如聚苯乙烯滴直径)与内置的荧光染料的波长参数从而获得不同发射波长的激光信号。
161、6.3.3荧光蛋白的表达
162、基于源自水母细胞的绿色荧光蛋白基因序列,多核苷酸编码可以在靶神经解剖结构的神经元中表达光响应性视蛋白。先利用基因工程技术(包括表达载体的构建与转染)让发光生物体的细胞表达绿色荧光蛋白(包括透皮方案、可植入方案、使用多种照射波长的方案、脉冲方案、组织界面等),然后让表达荧光蛋白的细胞借助镜子的反射制造激光信号。激光照射靶组织结构将刺激光敏蛋白发光。靶组织结构可以选自:脊髓、神经细胞体、神经节、神经纤维或神经末梢。激光照射装置包含光递送元件与传感器(包括但不限于:光学传感器、温度传感器、化学传感器和电传感器);输出信号参数涵盖:电流、电压、光功率(opticalpower)、辐照度、脉冲持续时间、脉冲间隔时间、脉冲重复频率和占空比(dutycycle)。电流控制器的变量包括了电流幅值、脉冲持续时间、占空比和递送的总体能量。电流脉冲的持续时间在1毫秒至100秒,占空比在99%至0.1%。
163、将光响应性视蛋白引入细胞的方法包括病毒介导的基因递送、电穿孔、光穿孔(optoporation)等。光敏蛋白可以是视蛋白蛋白质(例如,去极化视蛋白、超极化视蛋白、刺激性视蛋白、抑制性视蛋白、嵌合视蛋白)。转录启动子可以选自hsyn、cmv、hb9hb和ef1a。病毒构建体可以选自:aav5-hsyn-enphr3.0、aav6-hsyn-enphr3.0、aav6-cag-enphr3.0、aav8-hsyn-enphr3.0、aav8-hsyn-ic1c23.0。光源可以配置成发射波长在选自以下的波长范围内的光:440至600nm、601至700nm。光递送元件可以包括发光二极管(led)。光递送元件可以放置在神经或神经束圆周的大约60%处。可使用显像设备(如腹腔镜相机、超声导管、荧光镜等)。将外科手术装置(相机、针、工具等)插入皮肤的小切口位置,由定位设备指向靶组织。对于实验动物(例如小鼠),选择神经束、被神经纤维包绕的血管、或需要注射的其他稍圆柱状的靶解剖结构,注射载体溶液或凝胶。可以可控地增加注射器内的压力(例如,使用输注泵或注射器),以向解剖结构注射所需的溶液或凝胶。
164、6.3.4重组病毒载体递送
165、本发明建议采用重组病毒载体递送视蛋白与启动子dna序列,涉及转染靶神经并诱导神经元产生光敏蛋白。病毒可以递送至不同于靶组织结构的解剖位置,递送方式涉及注射、输注或滴注。将多核苷酸递送至靶神经解剖结构后需要确认靶神经表达了光响应性视蛋白(等待时长在1至6个月之间)。载体溶液可以扩散至整个组织并被细胞体吸收(100至1000个细胞)。携带视蛋白的病毒载体涵盖视蛋白(例如chr2、nphr等)和选择性启动子。慢病毒可以使用标准组织培养和超离心技术容易地生产,而aav可以可靠地由各单独的实验室或通过核心病毒机构产生。慢病毒和腺相关病毒(“aav”)载体已经成功地被用于将视蛋白引入小鼠、大鼠和灵长类动物脑中。
166、可以施用溶液或凝胶来感染皮肤的靶神经纤维,也可以使用微穿刺设备引入病毒载体。
167、6.4关于发光生物体激光谐振腔运行机制的数学模型:
168、下边先采用数学模型介绍激光谐振腔的稳定性条件,然后陈述所述纳米尺度的激光谐振腔如何调控参数从而满足激光谐振腔的稳定性条件。以聚苯乙烯滴为例的纳米尺度的激光谐振腔具有液滴形状,为便于建立数学模型,对应的激光谐振过程可以被简化为在两个反射镜之间出现激光谐振。本文假设介质充满了位于两个反射镜之间的空间。
169、所述纳米尺度的激光谐振腔,基本特性在于满足激光谐振腔的稳定性条件并产生激光谐振。
170、6.4.1激光谐振腔稳定发光的必要性条件包含“信号正反馈条件”与“增益系数超出临界值”。“增益系数超出临界值”中临界值受制于光能损耗与两个发射镜的透射率。激光谐振腔的“相位滞后整数倍条件”指腔内往返一周的相位滞后为2π的整数倍(光的初始位置可以位于任何一个相位点)。激光谐振腔的“信号正反馈条件”指腔长等于半波长的整数倍,意味着腔内形成稳定的驻波图案(波峰数目为整数)。
171、从能量角度看,激光谐振腔的“信号正反馈条件”要求腔长等于半波长的整数倍(腔内形成稳定的驻波图案),体现出系统的总能量维持在一个恒定的数值。为实现这一目标,从增益介质获取的能量必须等于损耗的能量。在物理学中通过向机械振子(例如单摆)补充机械能以维持恒定机械振幅,向电磁场补充电磁能以维持恒定电磁波振幅。
172、考虑到增益介质存在饱和效应(大信号输入会降低增益介质的效率),从增益介质获取的能量必须稍稍大于损耗的能量(能量损耗过程发生在腔内传输过程中以及在两个反射镜的透射过程中)。可以将激光谐振腔看作是曲率半径r1、r2的反射镜,或焦距f1=r1/2、f2=r2/2的透镜(透镜间距离等于相应腔镜之间的距离)。
173、采用a(i,:)表示a矩阵第i行的全部元素,则已知曲率半径为r的凹面反射镜的光束变换矩阵a为:2*2矩阵a中a(1,:)为[1,0],a(2,:)为[-2/r,1]。
174、考虑谐振腔内包含了曲率半径r1、r2的反射镜,则光线在谐振腔内往返一周的光束变换矩阵b为:b=a1*c*a2*c,其中2*2矩阵a1中a1(1,:)为[1,0],a1(2,:)为[-2/r1,1];2*2矩阵c中c(1,:)为[1,l],c(2,:)为[0,1];2*2矩阵a2中a2(1,:)为[1,0],a2(2,:)为[-2/r2,1]。矩阵a1对应着曲率半径r1的反射镜,矩阵a1对应着曲率半径r2的反射镜。
175、采用微积分方法建立如下的数学模型:
176、入射光强为iin,光离开一个端面(反射镜)进入增益介质中,在介质中的光速必定会小于真空中光速c,对应的能量损耗将大于在真空中传播的情形。
177、为简化讨论,本文认为在介质中的光速近似地等于c。单色光的光强正比于光子密度,于是i=hνcn(ν为光子频率)。
178、将z方向定义成沿着光传播方向,于是,(z,z+dz)区间对应着在某个点z对应的微分区间长度dz。光子从某个点z传播至(z,z+dz)区间的末端位置(z+dz)所消耗的时间为dt=dz/c,i(z+dz)-i(z)=dz[i(z)σ21(ν)(n2(z)-n1(z))]=dz[i(z)σ21(ν)*δ(n(z))]此处δ(n(z))=(n2(z)-n1(z))对应着高能级原子数n2(z)减去低能级原子数n1(z)。
179、当微分dz趋近于零时,差分i(z+dz)-i(z)可以用di(z)来表示,于是
180、di(z)=dz[i(z)σ21(ν)*δ(n(z))],处于发光细胞中的纳米尺度激光谐振腔内的光强很弱,于是任意时刻的原子数量的变化可以近似地等于初始时刻状态。δ(n(z))对应于某个微分区间(z,z+dz)的原子数量的变化值,它等于初始时刻(初始位置z=0)的原子数量的变化值δ(n(0))=δn0。δn0指初始时刻的原子数量的变化值(高能级原子数量与低能级原子数量之间的差值)。σ21(ν)指与入射光频率ν相关的一个常数(σ21的下标2对应着高能级原子数,下标1对应着低能级原子数)。
181、6.4.2光强的变化过程符合指数函数规律:
182、i(z)=iinexp[z*σ21(ν)*δn0],以iin作为指前系数,而指数部分z指沿着光传播方向的某个位置点,σ21(ν)指与入射光频率ν相关的一个常数,δn0指初始时刻的原子数量的变化值(高能级原子数量与低能级原子数量之间的差值)。光强正比于电磁振幅绝对值平方。设反射镜m1与m2的振幅透射率分别为r1与r2;振幅反射率分别为t1与t2。
183、可以把激光器的一个反射镜作全发射镜(光强反射率r(1)=1),另外一个反射镜作部分透射镜。光强反射率r(1)=(r1)2;r(2)=(r2)2。增益系数g0对应于σ21(ν)*δn0。初始时刻从反射镜m1入射的光是振幅能量为e0的平面波,增益介质的长度为l,从反射镜m2透射,e(0)=e0t1t2exp[(g0-δ)l/2-ikl]。在腔内往返一周后,从反射镜m2透射,则e(1)=e0*r1*r2*t1t2exp[(g0-δ)3l/2-i3kl];e(2)=e0(r12)*(r22)*t1*t2*exp[5*(g0-δ)l/2-5ikl];e(3)=e0(r13)*(r23)*t1*t2*exp[7*(g0-δ)l/2-7ikl];更普遍意义的表达式为e(n)=e0(r1n)*(r2n)*t1*t2*exp{(2n+1)*[(g0-δ)l/2-ikl]}。
184、多光束干涉的累积效应:∑e(n)=∑e0(r1n)*(r2n)*t1*t2*exp{(2n+1)*[(g0-δ)l/2-ikl]}=(e0*t1*t2)*∑(r1n)*(r2n)*exp{(2n+1)*[(g0-δ)l/2-ikl]}=(e0*t1*t2)*{exp[(g0-δ)l/2-ikl]}/{1-r1*r2*exp[(g0-δ)l-2ikl]}。∑e(n)与e0的比值趋向无穷大∞,即∑e(n)趋向无穷大∞,于是∑e(n)的分母趋向于零,意味着{1-r1*r2*exp[(g0-δ)l-2ikl]}的实部与虚部趋近于零。
185、6.4.3激光谐振腔稳定发光的必要性条件包含了“相位滞后整数倍”与“稳定驻波的能量补偿”条件。
186、虚部为零对应着激光谐振腔的“相位滞后整数倍条件”。2kl=2πq(q为大于或等于零的整数),意味着平面波在腔内往返一周,出发位置(初始位置)可以是任意的一个数值,相位滞后是2π的整数倍。腔长等于半波长的整数倍指l=q*(λq/2),其中l为腔的长度(两镜之间的距离),λq/2指半波长,q是大于等于零的整数。
187、实部为零对应着激光谐振腔的“稳定驻波的能量补偿”条件。从增益介质获得的能量补充必需足以弥补能量损耗(考虑到增益介质的效率会出现稍稍下降,从增益介质获得的能量需要稍稍超出能量损耗的数值)。由1-(r1*r2)exp(2g0-2δ)=0得到exp(2g0-2δ)=1/(r1*r2)将等式右边变成exp[-ln(r1*r2)],于是2g0-2δ=-ln(r1*r2)。
188、上述研究中稳定驻波的能量补偿条件指出了必不可少的能量条件。如果希望实现自激振荡,除了满足上述的能量补偿条件,还需要确保超出最低能量门槛(提供能量补偿以维持正反馈机制,并且由稳定驻波体现了足够的能量放大倍数)。
189、6.5关于发光生物体激光谐振腔的质量控制体系
190、考虑到发光生物体的激光谐振腔存在许多不确定因素,现实世界中难以准确判断是否满足激光谐振腔的稳定性条件(“信号正反馈条件”、“增益系数超出临界值”、“相位滞后整数倍”与“稳定驻波的能量补偿”条件)。本发明通过质量控制体系确保生物光子智能发射器具有“信号强度稳定”与“噪音信号极弱”等特点。
191、本发明采用数学模型阐释了激光谐振腔的稳定性条件,有益效果是,利于通过参数指标来评判生物光子智能发射器的运行效率。优选地,采用“增益系数”、“相位滞后”、“能量补偿临界值”等参数指标可以优化激光谐振腔从而满足激光谐振腔的稳定性条件。本发明的特点是以理论推导的结果(激光谐振腔的稳定性条件)来指导生物光子智能发射器中纳米尺度的激光谐振腔的设计制造与运行。激光谐振腔的稳定性条件属于生物光子发射器的信号输出效果,纳米尺度的激光谐振腔属于信号输出效果的内在制约因素与结构基础。本发明的技术策略是通过评判结果的优劣来优化设计制造与运行,这是以结果为导向来优化生产过程。
本文地址:https://www.jishuxx.com/zhuanli/20240802/235514.html
版权声明:本文内容由互联网用户自发贡献,该文观点仅代表作者本人。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如发现本站有涉嫌抄袭侵权/违法违规的内容, 请发送邮件至 YYfuon@163.com 举报,一经查实,本站将立刻删除。
下一篇
返回列表