一种真菌源昆虫蜕皮激素UDP糖基转移酶及其过表达载体构建方法与应用
- 国知局
- 2024-06-20 11:38:59
本发明涉及生物工程,具体涉及一种真菌源昆虫蜕皮激素udp糖基转移酶及其过表达载体构建方法与应用。
背景技术:
1、生物农药具有来源广泛、对靶标害虫特异性强、不易杀伤非靶标和有益生物、防治效果持久、对环境友好、害虫不易产生抗药性等优点,是病虫害综合治理(ipm)的关键组成部分(张慧等,2023;lacey et al.,2015)。真菌类杀虫剂是生物农药产业的重要组成部分,是一类寄主谱较广的病原真菌制剂,可通过直接穿透昆虫的角质层来感染昆虫,不仅可以控制咀嚼式害虫,而且对具有吸吮口器的昆虫,如蚜虫和蚊子等也有很好地杀灭效果。昆虫病原真菌高度特异性,对环境安全,且通常具有较高的毒力,能导致大规模的昆虫流行病,因此非常适合触杀类真菌杀虫剂的开发(李增智,2015;wang et al.,2015;wang&wang,2017)。
2、迄今已有50多种登记注册的真菌杀虫剂,包括球孢白僵菌、金龟子绿僵菌、玫烟色拟青霉和蜡蚧轮枝菌在内的十多种真菌(mazid et al.,2011;lacey et al.,2015)。与其他类生物杀虫剂相比,杀虫真菌对寄主的侵染机理和过程较为独特和复杂,是病原物和寄主之间相互生理生化作用的结果。以绿僵菌(metarhizium)为代表的子囊菌(在生活史上不存在有性阶段)为例,关键侵染阶段包括:感染繁殖体(例如分生孢子和芽孢子)粘附到宿主角质层的表面;感染结构(附着胞和穿透针)的分化;使用酶和机械压力穿透角质层;血体腔的定植,克服昆虫免疫,然后杀死寄主;分生孢子在虫体外部出现并开始下一轮侵染(buttet al.,2016;wang&wang,2017)。另一方面,因为真菌杀虫过程复杂而精细,所以真菌杀虫剂也具有杀死寄主昆虫速度慢,对环境条件敏感等缺点,严重制约了它农药市场的应用及推广(hong et al.,2023)。因此,真菌侵染致病性和抗逆性相关功能基因的发掘、并利用它们进行生产菌株遗传改良的生物技术,为开发杀虫效果更好、环境稳定性更强的真菌杀虫刹、杀菌剂是目前生物防治技术发展的重要方向之一。
3、真菌毒力和抗逆性相关功能基因的发掘体围绕以上侵染的全过程展开,其中真菌在昆虫体内定殖过程包含了许多适应性策略和具有潜力的相关功能酶和基因,值得深入探明和挖掘。杀虫真菌进入血体腔后能够与昆虫的免疫系统相互作用并成功感染寄主,这其中包括干扰、破坏或操纵基础免疫或诱导型免疫等免疫逃避策略,是昆虫病原真菌与昆虫的免疫机制进行动态干扰的过程(lu et al.,2015)。有研究表明,昆虫体内的激素也是影响昆虫病原真菌成功定殖的重要因素(kamimura et al.,2012;will et al.,2020),因为昆虫激素不仅参与和控制昆虫蜕皮变态的过程,还很大程度上参与调控昆虫免疫反应(liuet al.,2014)。如蜕皮激素在昆虫胚胎发育各个时期的滴度均有波动,尤其是在中期有一个滴度高峰(thummel,2001)。如球孢白僵菌接种大蜡螟幼虫后,幼虫体内血淋巴的蜕皮激素(20e)浓度逐步上调,在第4d高于对照组32.9%,说明20e参与了侵染过程中病原真菌与宿主昆虫的互作(冯雪瑶,2017)。越来越多的研究表明蜕皮激素主要影响昆虫的细胞免疫。使用章胺、5-羟基色胺或多巴胺处理蟑螂后,其体内血淋巴中的吞噬细胞活性增加(baineset al.,1992)。类似地,蜕皮激素能增强黑腹果蝇的吞噬作用,可以增强结节的形成,这可能与蜕皮激素可诱导细胞分化成巨噬细胞有关(dimarcq et al.,1997;franssens etal.,2006)。当受到细菌感染时,果蝇唾液腺中类固醇蜕皮激素的转录激活与双翅肽基因的免疫应答反应一致,蜕皮激素对果蝇的免疫应答反应具有促进作用(meister et al.,1996)。蜕皮激素20e可促进与昆虫先天免疫相关的habbp蛋白显著上调表达(wang et al.,2011);而其在鳞翅目昆虫中可以激活抑血细胞聚集素的显著上调表达,以调节血细胞对病原菌的黏附(shaik and sehnal,2009);20e还能增强昆虫细胞间的粘连作用,使吞噬率提高约15%,对细胞免疫有明显增强作用(柴连琴,2015);故通过失活昆虫体内20e能有效提高球孢白僵菌对寄主的侵染效率(冯雪瑶,2017)。
4、昆虫病原菌与寄主昆虫的共同进化过程中,有些病原菌在与寄主相互作用过程中适应性地进化过程中产生的酶出了能特异性地与蜕皮激素作用使蜕皮激素失活的酶,从而达到减弱昆虫免疫反应、改变寄主行为并致死的效果。目前,除了c22位羟基氧化酶在昆虫病原真菌中已有报道外,其余酶绝大多数报道来自昆虫,少数见于昆虫病毒和细菌。研究较多的是昆虫杆状病毒,其编码的蜕皮类固醇udp-葡糖基转移酶(egt),能通过糖与蜕皮酮c22位结合位使之失活,进而阻止幼虫的蜕皮、影响昆虫的生长发育,使蜕皮激素失活,阻止幼虫的蜕皮(clarke et al.,1996)。昆虫病原真菌中也存在类似的蜕皮激素失活酶——ecdysteroid 22-kinase(eckinase),能有助于真菌调控昆虫体内的昆虫激素20e的浓度,从而降低昆虫体内的免疫水平,有助于真菌快速战胜寄主免疫,加快对寄主的击倒速度。目前已经被证实的报道的有莱氏野村菌(nomuraea rileyi)中的eckinase(mre22o)能特异性将蜕皮酮c22位上的羟基氧化成羰基,使蜕皮酮失去活性(kiuchi et al.,2003)。目前,已有研究将这类蜕皮激素失活酶基因从昆虫病毒或真菌中克隆重组至球孢白僵菌(beauveria bassiana)中,使之表达量升高,从而达到降低昆虫体内20e浓度、降低昆虫免疫反应,快速杀虫的目的。如将来自加利福尼亚核多面体病毒(acmnpv)的昆虫蜕皮激素udp糖基转移酶基因(egt)重组转化球孢白僵菌,该基因能使昆虫发育中的关键激素20-羟基蜕皮激素失活,可提高球孢白僵菌感染早期家蚕幼虫的死亡率(mao et al.,2023)。将莱氏野村菌(nomuraea rileyi)中的eckinase(mre22o)基因重组至球孢白僵菌(beauveriabassiana)中进行过表达后,能有效降低昆虫体内20e的浓度,抑制昆虫的免疫反应,使得真菌杀虫速度和活性得到提升(zhu et al.,2021)。
5、然而,目前除了莱氏野村菌中的蜕皮激素失活酶eckinase,暂未有相关专利或文献在昆虫病原真菌中鉴定出蜕皮激素udp糖基转移酶相关的基因。
6、因此,在被毛孢属专性真菌或是其他广谱性的昆虫病原真菌中高表达这个糖基转移酶,其在侵染宿主的过程中可能降低昆虫体内20e水平并抑制昆虫的免疫反应,将会有助于真菌缩短击倒宿主时间,提高真菌杀虫剂的杀虫活性,解决这类真菌杀虫活性不高的问题。
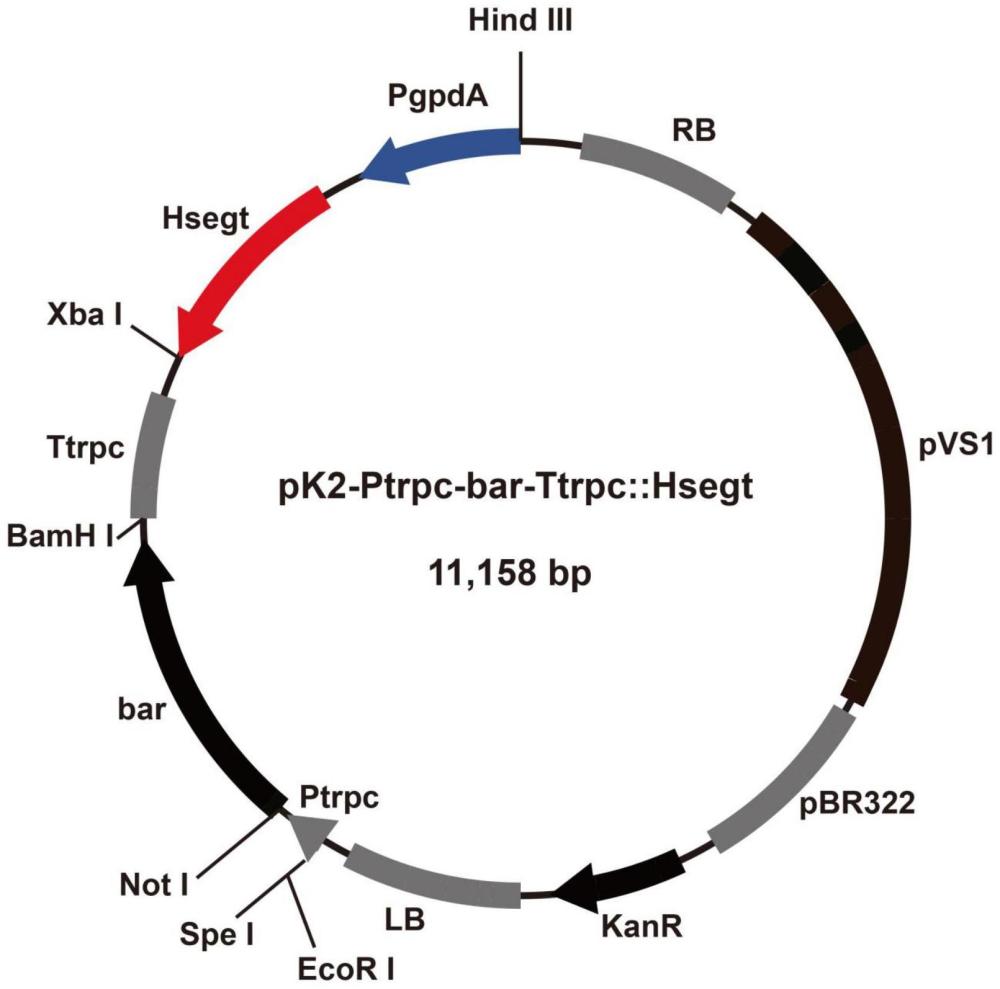
7、参考文献
8、(1)mitchell,c.,hawes,c.,iannetta,p.,birch,a.n.e.,begg,g.,&karley,a.j.anagroecological approach for weed,pest and disease management in rubusplantations.inraspberry).springer,2018.cham.pp.63–81.
9、(2)张慧,许宁,曹丽茹,王蕊,张凯.我国微生物农药的研发与应用研究进展.
10、农药学学报,2023,25(4):769-778.
11、(3)lacey,l.a.,grzywacz,d.,shapiro-ilan,d.i.,frutos,r.,brownbridge,m.&
12、goettel,m.s.insect pathogens as biological control agents:back to thefuture.journal ofinvertebrate pathology.2015,132:1–41.
13、(4)李增智.我国利用真菌防治害虫的历史,进展及现状.中国生物防治学报.
14、2015,31(5):699-711.
15、(5)wang,j.j.,qiu,l.,cai,q.,ying,s.h.&feng,m.g.transcriptional controloffungal cell cycle and cellular events by fkh2,a forkhead transcriptionfactor in an insectpathogen.scientific reports.2015,5:10108.
16、(6)wang,c.,&wang,s.insect pathogenic fungi:genomics,molecularinteractions,and genetic improvements.annual review of entomology.2017,62:73–90.
17、(7)mazid s,kalida jc,rajkhowa rc.a review on the use of biopesticidesin insect pest management.international journal of science and advancedtechnology.2011,1(7):169–178.
18、(8)butt,t.m.,coates,c.j.,dubovskiy,i.m.&ratcliffe,n.a.chapter nine-entomopathogenic fungi:new insights into host–pathogen interactions.advancesin genetics.2016,94:307–364.
19、(9)hong s,shang j,sun y,tang g,wang c.fungal infection of insects:molecular insights and prospects.trends in microbiology.2023,32(3):302-316.
20、(10)lu,h.l.,wang,j.b.,brown,m.a.,euerle,c.&st leger,r.j.identification of drosophila mutants affecting defense to anentomopathogenic fungus.scientific reports.2015,5:12350.
21、(11)kamimura,m.,saito,h.,niwa,r.,niimi,t.,toyoda,k.,ueno,c.&kiuchi,m.fungal ecdysteroid-22-oxidase,a new tool for manipulating ecdysteroidsignaling and insect development.journal of biological chemistry.2012,287(20):16488–16498.
22、(12)will i,das b,trinh t,brachmann a,ohm ra,de bekker c.geneticunderpinnings of host manipulation by ophiocordyceps as revealed bycomparative transcriptomics.g3(bethesda).2020,10(7):2275-2296.
23、(13)liu,w.,xie,y.,dong,j.,xue,j.,zhang,y.,lu,y.&wu,j.pathogenicity ofthree entomopathogenic fungi to matsucoccus matsumurae.plos one.2014,9(7):e103350.
24、(14)thummel,c.s.molecular mechanisms of developmental timing inc.elegans and drosophila.developmental cell.2001,1(4):453–465.
25、(15)冯雪瑶.通过失活昆虫蜕皮激素20e提高球孢白僵菌的侵染效率.西南大学.2017,中国,重庆.
26、(16)baines,d.,desantis,t.&downer,r.g.octopamine and 5-hydroxytryptamine enhance the phagocytic and nodule formation activities ofcockroach(periplaneta americana)haemocytes.journal of insect physiology.1992,38(11):905–914.
27、(17)dimarcq,j.l.,imler,j.l.,lanot,r.,ezekowitz,r.a.b.,hoffmann,j.a.,janeway,c.a.&lagueux,m.treatment of l(2)mbn drosophila tumorous blood cellswith the steroid hormone ecdysone amplifies the inducibility of antimicrobialpeptide gene expression.insect biochemistry and molecular biology.1997,27(10):877–886.
28、(18)franssens,v.,smagghe,g.,simonet,g.,claeys,i.,breugelmans,b.,deloof,a.&broeck,j.v.20-hydroxyecdysone and juvenile hormone regulate thelaminarin-induced nodulation reaction in larvae of the flesh fly,neobellieriabullata.developmental&comparative immunology.2006,30(9):735–740.
29、(19)meister,m.&richards,g.ecdysone and insect immunity:the maturationof the inducibility of the diptericin gene in drosophila larvae.insectbiochemistry and molecular biology.1996,26(2):155–160.
30、(20)wang,g.,liu,p.c.,wang,j.x.,&zhao,x.f.a btb domain-containing geneis upregulated by immune challenge.archives of insect biochemistry andphysiology.2011,77(2):58–71.
31、(21)shaik ha,sehnal f.hemolin expression in the silk glands ofgalleria mellonella in response to bacterial challenge and prior to celldisintegration.journal of insect physiology.2009,55(9):781-7.
32、(22)柴连琴,苗迎春,王乐.20-羟基蜕皮酮对棉铃虫免疫系统的影响.四川动物.2015,(5):739-747.
33、(23)clarke,e.e.,tristem,m.,cory,j.s.,&o'reilly,d.characterization ofthe ecdysteroid udp-glucosyltransferase gene from mamestra brassicaenucleopolyhedrovirus.the journal of general virology,1996,77(11):65-71.
34、(24)kiuchi,m.,yasui,h.,hayasaka,s.&kamimura,m.entomogenous fungusnomuraea rileyiinhibits host insect molting by c22-oxidizing inactivation ofhemolymph ecdysteroids.archives of insect biochemistry and physiology.2003,52(1):35–44.
35、(25)mao x,xing d,liu d,xu h,hou l,lin p,xia q,lin y,sheng.ecdysteroid udp-glucosyltransferase expression in beauveria bassianaincreases its pathogenicity against early instar silkworm larvae.journal offungi.2023,9(10):987.
36、(26)zhu s,feng x,keyhani no,liu y,jin d,tong s,pei y,fany.manipulation of host ecdysteroid hormone levels facilitates infection bythe fungal insect pathogen,metarhizium rileyi.environment microbiology.2021,23(9):5087-5101.
技术实现思路
1、针对现有真菌杀虫剂杀虫速度慢,特别是专性杀虫真菌杀虫活性不高的问题,本发明提供的一种真菌源昆虫蜕皮激素udp糖基转移酶及其过表达载体构建方法与应用,旨在解决上述背景技术中存在的问题,本发明提供了一种新的真菌源的蜕皮类固醇尿苷5’-二磷酸-葡糖基转移酶hsegt,具有以昆虫蜕皮激素20e为底物,在其分子c22位上添加糖基使20e失活,从而降低20e水平的功能。将其应用于杀虫真菌剂,能干扰宿主昆虫的相关激素水平,过表达降低昆虫的免疫抵御能力,以此改良专性杀虫真菌鹿儿岛被毛孢的杀虫效力,解决该类真菌侵染效率低的问题。
2、为了实现上述技术目的,本发明主要采用如下技术方案:
3、本发明公开了一种真菌源昆虫蜕皮激素udp糖基转移酶hsegt,具有如seq idno.2所示的氨基酸序列。
4、>hsegt seq id no.2
5、
6、本发明还公开了一种编码如上所述的真菌源昆虫蜕皮激素udp糖基转移酶hsegt的基因,具有如seq id no.1所示的核苷酸序列。
7、>hsegt seq id no.1
8、
9、
10、
11、本发明还公开了一种真菌源昆虫蜕皮激素udp糖基转移酶hsegt的过表达载体,为hsegt与来自构巢曲霉的三磷酸甘油醛脱氢酶编码基因的组成型启动子pgpda链接,组成hs::egt片段后,与过表达载体ppk2-bar连接,得到ppk2-bar-pgpda-hs::egt重组过表达载体。
12、本发明还公开了一种如上所述的过表达载体的构建方法,包括如下步骤:
13、(1)hsegt基因的克隆:设计引物,以杀虫真菌鹿儿岛被毛孢(hirsutellasatumaensis)的基因组dna为模版,扩增hsegt基因的全长编码序列,在hsegt基因的3’端添加his标签cat cat cac cat cac cat;同时在hsegt基因的5’端添加启动子gpda片段的3’端一段同源片段;在hsegt基因的3’端添加ppk2-bar质粒hindⅲ酶切位点上游的一段同源片段;
14、上游引物hsegt-f:ccgtcgacctcgactctagaatgtggttaaagtttttcta,如seq idno.3所示;
15、下游引物hsegt-r:acgacggccagtgccaagcttttagtggtgatggtgatgatgcaaagccttgaattttc,如seq id no.4所示;
16、(2)启动子pgpda片段的克隆:设计引物,以构巢曲霉(aspergillus nidulans,fgsc a4)基因组为模版,扩增得到启动子pgpda片段,并在pgpda片段的5’添加ppk2-bar质粒xba i酶切位点上游的一段同源片段;
17、上游引物pgpda-f:gaggtaatccttctttctagatgcaaagcatgcggagaga,如seq idno.5所示;
18、下游引物pgpda-r:tctagagtcgaggtcgacgg,如seq id no.6所示;
19、(3)重组质粒的合成:先分别克隆出hsegt基因片段和pgpda片段,将ppk2-bar质粒使用xba i和hindⅲ两种限制性内切酶酶切线性化;然后将这三种片段使用genearttmgibson assembly hifi克隆试剂盒连接起来,将hsegt基因片段置于pgpda启动子下游,测序验证后获得重组质粒ppk2-bar-pgpda-hs::egt;
20、(4)将重组质粒ppk2-bar-pgpda-hs::egt转入宿主菌感受态细胞中,设计验证引物进行pcr筛选和质粒的测序验证,获得ppk2-bar-pgpda-hs::egt重组过表达载体;
21、上游引物ex-ps-f:tgcaaagcatgcggagaga,如seq id no.7所示;
22、下游引物ex-ps-r:ttagtggtgatggtgatgatgcaaagccttgaattttc,如seq id no.8所示。
23、在本发明的较佳实施方式中,步骤1中,所述杀虫真菌鹿儿岛被毛孢(hirsutellasatumaensis)可以被汤普森被毛孢(hirsutella thompsonii)、球孢白僵菌(beauveriabassiana)、金龟子绿僵菌(metarhizium anisopliae)、黄绿绿僵菌(metarhiziumflavoviride)、布氏白僵菌(beauveria brongniartii)、玫烟色拟青霉(paecilomycesfumosoroseus)中的任意一种代替。
24、在本发明的另一较佳实施方式中,步骤4中,所述宿主菌选自大肠杆菌dh5α、农杆菌(agrobacterium tumefaciens,agl-1)或鹿儿岛被毛孢(gzuifr-hir201012jc)中的任意一种。
25、本发明还公开了一种真菌源昆虫蜕皮激素udp糖基转移酶hsegt的过表达工程菌株,将如上所述的过表达载体ppk2-bar-pgpda-hs::egt通过农杆菌agl-1介导方法导入鹿儿岛被毛孢菌丝中,获得hsegt基因过表达的重组工程菌株。
26、本发明还公开了如上所述的过表达工程菌株在制备杀虫真菌剂中的应用。
27、本发明还公开了一种杀虫真菌剂,含有如上所述的真菌源昆虫蜕皮激素udp糖基转移酶hsegt或如上所述的过表达载体或如上所述的过表达工程菌株,所述杀虫真菌剂使得昆虫体内的蜕皮激素20e失活、浓度下降、抑制寄主体内酚氧化酶活性及促进真菌虫菌体在寄主体内的增殖速率。
28、与现有技术相比,本发明具有如下有益效果:
29、目前,商品化的真菌杀虫剂主要是以球孢白僵菌、金龟子绿僵菌为代表的寄主范围较广泛的昆虫病原真菌,多为分生孢子粉剂,产品多样性不高,还没有针对特定寄主害虫的杀虫真菌。另外,白僵菌等寄主范围较广的真菌在田间应用时也较容易出现寄主产生抗性现象,需要有特异性强、杀虫作用方式独特的专性杀虫真菌产品。因此,针对该现象,从昆虫病原真菌中发现一种蜕皮类固醇尿苷5’-二磷酸-葡糖基转移酶hsegt,与昆虫及昆虫杆状病毒中报道过的egt高度同源,且具有蜕皮激素失活活性。将其在鹿儿岛被毛孢中进行过表达改良,增加其在该真菌中的基因表达量以及酶的分泌,有效提高该真菌对寄主昆虫的击倒速度,有效增加杀虫活性。以解决杀虫真菌田间应用时存在的作用时间长、活性不高的缺点。
30、主要表现在以下几点:
31、1、目前该类蜕皮类固醇尿苷5’-二磷酸-葡糖基转移酶一般来自昆虫或昆虫杆状病毒,而在昆虫病原真菌中该类蜕皮激素失活酶尚未见报道。本发明首次从真菌中发现的新的egt酶,这对真菌的毒力的以及对昆虫的侵染能力有帮助;
32、2、通过强启动子的融合,可以极大地提高该真菌中该蜕皮类固醇尿苷5’-二磷酸-葡糖基转移酶在真菌中的基因表达量以及酶的分泌,有效提高该真菌对昆虫体内蜕皮激素的失活,减少蜕皮激素浓度,进而降低相应的昆虫免疫反应,促使真菌快速生长,杀死昆虫;
33、3、本发明得到的改良的工程菌株增强了蜕皮类固醇尿苷5’-二磷酸-葡糖基转移酶的表达量,能有效缩短杀死寄主昆虫的时间。
本文地址:https://www.jishuxx.com/zhuanli/20240619/1813.html
版权声明:本文内容由互联网用户自发贡献,该文观点仅代表作者本人。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如发现本站有涉嫌抄袭侵权/违法违规的内容, 请发送邮件至 YYfuon@163.com 举报,一经查实,本站将立刻删除。
下一篇
返回列表