用于增强抗体可变区基因突变频率的方法、抗体可变区基因突变方法及其应用
- 国知局
- 2024-09-14 14:23:51
本发明属于生物医药领域,特别是涉及用于增强抗体可变区基因突变频率的方法、抗体可变区基因突变方法及其应用。
背景技术:
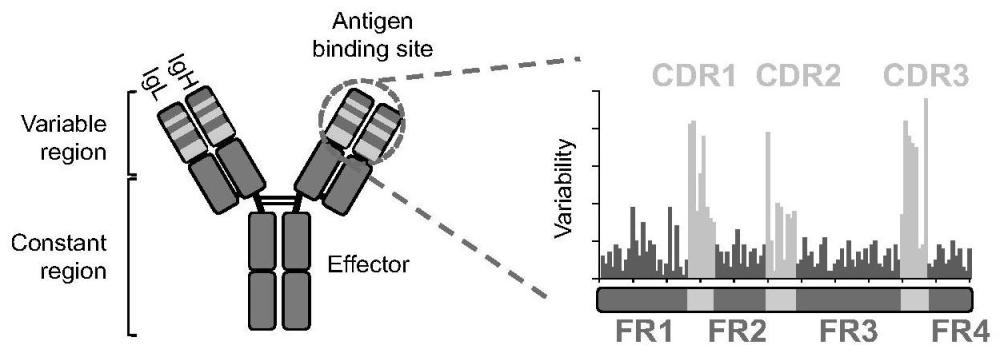
1、抗体的结构类似一个y字形,由两条相同的重链和两条相同的轻链组成。抗体的n端负责结合抗原,多样化非常高,故称为可变区(variable region)。可变区由间隔的框架区(framework region,fr)和互补决定区(complementarity determining region,cdr)组成,其中fr负责维持抗体的结构,序列较为保守,而cdr负责结合抗原,序列多样性非常高。抗体的c端序列相对保守,称为恒定区(constant region),决定抗体的类型并负责下游信号的传导(图1左图)。
2、哺乳动物的抗体多样化过程主要分为两个阶段,其分子机制和功能各不相同。在抗原暴露之前,骨髓中的b细胞通过rag内切酶介导的v(d)j重排将重链和轻链胚系基因的可变区(variable,v)、多样区(diversity,d)和连接区(joining,j)特异性重组形成初始抗体库1,2。然而,初始抗体库的抗原亲和力较低,并不能提供完善的免疫保护。为了丰富和提高初始抗体库的多样性及亲和力,外周淋巴器官中的b细胞需要进一步经历体细胞高频突变(somatic hypermutation,shm)和抗体类型转换(class switch recombination,csr)过程。shm过程在抗体可变区基因外显子中引入高频突变3,突变会产生不同亲和力的b细胞克隆,经过克隆选择,具有提高亲和力的克隆被保留,而带有有害突变的克隆则被淘汰4-6,这一使抗体逐渐增加亲和力的过程,被称为抗体亲和力成熟过程。亲和力成熟是体液免疫反应中产生针对入侵病原体高亲和力抗体的核心步骤。在抗体可变区基因外显子中,高频突变并非均匀分布,而是偏好积累在三个间隔排列的cdr区(图1右)7-9。cdr的高突变偏好似乎是固有存在的,因为无功能抗体基因小鼠模型在无抗原选择压力的情况下,依然表现出cdr的高突变偏好特征10-12。cdr和fr区域之间的密码子偏倚与这种突变偏好相关13-16。
3、抗体多样性的产生对机体抵御外界病原体入侵至关重要。在受到抗原刺激后,b淋巴细胞在其他免疫细胞的辅助下形成生发中心(germinal center,gc),并引发免疫反应。在生发中心(gc)b细胞中,激活诱导的胞苷脱氨酶(activation-induced cytidinedeaminase,aid)负责起始抗体的体细胞高频突变过程17,18。aid作用于抗体可变区基因,催化单链dna中的胞嘧啶核苷酸(c)脱氨转化为尿嘧啶核苷酸(u)。产生的u:g错配借助下游dna碱基切除修复(ber)和错配修复(mmr)过程引发多种类型的修复产物3,19,从而增加序列的多样性。aid20-22优先作用于wrcy(w=a/t、r=a/g和y=c/t)序列模块,下游跨损伤易错修复聚合酶(polη)23,24引入了进一步的突变,偏好作用于wa序列模块25。aid在可变区上引入的突变总是倾向于积累在cdr区而非fr区。aid如何特异地感知抗体可变区基因中的不同区域,一直是本领域内悬而未决的问题。体外实验中,重组aid蛋白优先将ssdna底物wrc模块中的c脱氨20-22,这与aid/apobec超家族中其他蛋白的微尺度序列偏好相似26。aid的微尺度脱氨酶序列模块(wrc)与cdr富含的agy(丝氨酸)密码子16,27相关,并且回文序列(如agct)中的重叠热点是b细胞中最常见的脱氨位点28,29。然而,脱氨酶序列模块本身无法解释cdr中的高突变偏好,因为wrc也存在于fr中,并广泛分布于全基因组范围。是否存在其他未知调控因素目前仍不得而知,也一直是领域内的研究热点之一。
4、在抗体可变区基因外显子区域内,固有shm图谱显示,与fr区相比,cdr区中的wrc序列模块发生更高频率的突变30。此外,定点敲入抗体可变区基因外显子位点的细菌和人源非抗体序列也表现出wrc序列模块突变频率的差异,这种特征类似于cdr与fr之间的偏倚12。这些发现表明,在中尺度水平(5-50bp)很可能存在某些未知且能够增强特定wrc序列模块突变频率的顺式元件。这一假设与ssdna底物必须具有一个最小长度才能有效脱氨的实验结果一致22。与抗体可变区基因外显子相反,哺乳动物抗体基因重链(igh)转换(s)区是csr过程中aid的主要靶标。s区内agct模块分布密集并且富含g串联重复序列31,这些序列特征通过形成复杂的二级结构促进aid脱氨32,33。然而,抗体可变区基因外显子和aid脱靶序列(off-targeting)34缺乏这些独特的dna特征,aid如何优先靶向cdr(或类cdr区域)中特定的wrc序列模块仍然是一个谜。
5、因此,在分子层面确定生物体如何操控aid优先靶向cdr(或类cdr区域)中特定的wrc,从而增强抗体可变区基因突变频率,无论在理论研究和体内外的分子生物学实际操作上,均具有深刻的理论意义和现实意义。
技术实现思路
1、鉴于以上所述现有技术的缺点,本发明的目的在于提供一种增强抗体可变区基因突变频率的方法,所述方法包括以下步骤:将抗体可变区基因中至少部分的aid偏好作用模块的调控元件置换为嘧啶核苷酸比例相当于或高于嘌呤核苷酸比例的片段,所述调控元件为aid偏好作用模块的5’端和/或3’端各6~18个核苷酸的片段。本发明的用于增强抗体可变区基因突变频率的方法可以用于解决现有技术中通过增加aid靶向cdr(或类cdr区域)中特定的wrc的倾向性,从而增强抗体可变区基因突变频率的问题。
2、本发明还提供一种抗体可变区基因突变方法,所述抗体可变区基因突变方法包括以下步骤:(1)采用权利要求1~5任一权利要求所述的方法改造待突变抗体可变区基因;以及(2)使用aid酶作用于经步骤(1)改造的抗体可变区基因,获得突变的抗体可变区基因。
3、为实现上述目的及其他相关目的,本发明提供一种抗体可变区基因突变体多核苷酸或抗体可变区基因突变多核苷酸文库,采用所述的抗体可变区基因突变方法制备获得。
4、本发明还提供所述的抗体可变区基因突变体多核苷酸或抗体可变区基因突变多核苷酸文库在构建抗体人源化小鼠上的用途。
5、本发明还提供一种核酸构建体或核酸构建体文库,所述核酸构建体包含所述的抗体可变区基因突变体多核苷酸;所述核酸构建体文库包括所述的抗体可变区基因突变体多核苷酸文库。
6、本发明还提供所述的增强抗体可变区基因突变频率的方法或所述的抗体可变区基因突变方法用于抗体进化的用途。
7、本发明还提供一种抗体进化的方法,所述抗体进化的方法包括利用所述的增强抗体可变区基因突变频率的方法或所述的抗体可变区基因突变方法筛选性能优化的抗体。
8、本发明还提供一种用于增强抗体可变区基因突变频率的试剂盒,所述试剂盒包含作为调控元件的5’端和/或3’端各6~18个核苷酸的片段,所述片段的嘧啶核苷酸比例相当于或高于嘌呤核苷酸比例;aid酶,以及aid酶的工作溶液。
9、本发明还提供了所述的试剂盒用于增强抗体可变区基因突变频率的用途。
10、如上所述,本发明的用于增强抗体可变区基因突变频率的方法、抗体可变区基因突变方法及其应用,具有以下有益效果:
11、(1)能显著提高特定序列(agct序列)的突变频率,和/或非wrc模块中的c碱基的突变频率,实现了在分子层面操作经脱氨导致突变,为进一步研究抗体多样化提供了基础;
12、(2)本发明提出的增强抗体可变区基因突变频率的方法、试剂盒和用途,为在体内外在分子层面操作wrc模块和非wrc模块的脱氨产生的突变,提供了新的分子生物学工具。
本文地址:https://www.jishuxx.com/zhuanli/20240914/293831.html
版权声明:本文内容由互联网用户自发贡献,该文观点仅代表作者本人。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如发现本站有涉嫌抄袭侵权/违法违规的内容, 请发送邮件至 YYfuon@163.com 举报,一经查实,本站将立刻删除。
下一篇
返回列表