GhVdR1基因在提高棉花黄萎病抗病性中的应用
- 国知局
- 2024-10-09 14:38:53
本发明属于生物技术应用领域,涉及一种棉花ghvdr1基因在提高棉花抗病性中的应用。该基因orf全长为1197bp,编码398个氨基酸。结构域分析发现其编码细胞色素b561蛋白(cyb561)。ghvdr1基因通过调控抗坏血酸再生在棉花中参与调控棉花抗病性。在棉花中该基因功能未有相关文献报道。本发明分析ghvdr1在陆地棉不同组织器官和受黄萎病菌诱导后不同时间点棉花根部中的表达水平,发现ghvdr1是棉花细胞色素b561家族基因中唯一一个在各组织中呈组成性表达且受黄萎病菌诱导后显著下调表达的基因。通过对不同植物中ghvdr1同源基因序列比对和进化分析,发现该基因结构在各个物种中非常保守。亚细胞定位结果证明ghvdr1主要定位于高尔基体膜,在内质网和细胞膜也有定位。在棉花中抑制ghvdr1基因表达,植株对黄萎病菌的抗性显著提高,而过量表达几乎无影响。进一步对rnai干扰植株进行转录组测序分析,与对照相比,ghvdr1转录水平的抑制影响了植株体内氧化还原反应相关通路和抗坏血酸再生途径关键基因的表达量。生化检测结果显示,干扰ghvdr1降低了植株中asa的含量和asa氧化还原效率,而h2o2含量显著增加。上述结果表明ghvdr1通过参与植株抗坏血酸再生来调节ros含量,影响棉花黄萎病抗性。外施asa降低了植株抗病性的实验结果为ghvdr1抗病机理解析提供进一步补充。转基因株系的产量和纤维品质相关性状表型调查结果显示,与受体相比,干扰ghvdr1对植株产量和纤维品质影响较小。本发明利用生物技术明确了ghvdr1调控棉花黄萎病抗病性的作用,提出通过抑制ghvdr1表达调控棉花抗病性的育种利用价值。
背景技术:
1、缺失免疫细胞的植物在应对复杂生存环境中的多种胁迫时不断进化,逐渐在体内形成由多种防御手段组成的较为完整的免疫体系,目前对植物免疫机制已经有了一定的研究。高等植物细胞表面和细胞内部拥有大量的免疫受体,可以感知病原菌感染相关的各种信号(kourelis and van der hoorn,2018;van de weyer et al.,2019)。细胞表面免疫受体称为模式识别受体(prrs),识别由病原菌释放的相对保守的微生物相关分子模式(mamps)和损伤相关分子模式(damps),例如:脂多糖、肽聚糖、真菌几丁质和源自植物的寡半乳糖醛苷等,启动pamps触发的免疫反应(pti)(zhu et al.,2022)。ve1是典型的prr,帮助番茄、拟南芥、烟草和棉花抗黄萎病(fradin et al.,2011;song et al.,2018);ve同源基因gbve1、gbavd1、gbavd2、gbvdr3、gbvdr5和gbvdr6同样正调控拟南芥和棉花抗黄萎病过程(li et al.,2015;chen et al.,2016;chen et al.,2017;yang et al.,2017)。在细胞内或跨膜的免疫受体是富含亮氨酸重复受体蛋白(nlr),通过特异性感知进入到植物细胞内的病原菌效应蛋白,启动效应器触发的免疫(eti)(ramirez-prado et al.,2018)。蛋白cnl、rpm1和rps2与质膜相关宿主蛋白rin4结合监测丁香假单胞菌效应子avrrpm1、avrb和avrrpt2,是nlr帮助植物抗病的经典例子(axtell and staskawicz,2003;mackey et al.,2002)。
2、pti和eti共同组成植物先天免疫系统,这一系统的工作模式最初被形象概括为“z模型”。pti作为第一层免疫反应发挥作用,部分病原菌为侵入植物细胞产生干扰pti扰乱植物免疫的效应因子,引起效应子触发的感病性(ets),植物细胞随即进化出能够识别效应因子的nlr,启动eti释放r蛋白阻止病原菌进一步扩繁(jones and dangl,2006)。植物在经典免疫模型中pti和eti一直被认为是相互独立的,但越来越多证据表明两者在信号级联和信号组分上存在潜在的联系。磷酸化蛋白质组学研究发现pti和eti的早期信号感知过程存在协同磷酸化现象(kadota et al.,2019);另一研究表明,在prr未被激活的情况下,激活nlr仅能诱导部分防御反应启动(ngou et al.,2020;yuan et al.,2020),而同时刺激prr和nlr才能完全激活防御,该实验还发现nlr的预激活也引起了prr介导的防御;还有研究表明两者在信号转导中会相互增强(ngou et al.,2020)。这些研究表明pti和eti之间存在信号串扰,具体原因还有待进一步研究。
3、尽管pti和eti的触发机制不同,但它们触发后启动的下游反应是相似的,包括:果胶积累、木质素和胼胝质沉积,抗菌毒素、酚类和黄酮类化合物等物质的积累,中继和放大下游信号的mapk信号级联途径,ca2+的内流,ros爆发,感知和传递信号的多种植物激素的生物合成,细胞程序性死亡(pcd)的发生,免疫相关基因的转录重编程,抗病基因的表达等(billah and yang,2021)。以上由病原菌激活的免疫反应可称为诱导免疫反应,系统获得性抗性(sar)是诱导免疫反应中的一种,指植株某部分产生对某种病原物的抗性后,将抗性传递给植株其他部位,使未受侵染的组织器官拥有同样或者更高的抗性。经研究,大多数sar关键信号分子是水杨酸(sa),当植株在自身体内积累到足够多sa,sar就会发生,ros也是启动sar必不可少的信号分子(王钧和金巧玲,1998)。由此可见,ros在植物免疫中非常重要。各种激素信号之间相互联系,相互作用,在正常野生型中研究十分困难,利用突变体可加速抗病机制研究进程。
4、ros在植物非生物和生物应激感应的初始阶段起重要作用,组织内ros的产生往往是植物受到胁迫的标志,而ros的产生也有利于植物建立后续防御,比如:改变细胞壁结构、启动激素信号转导通路、启动sar、诱发超敏反应(hr)等(qi et al.,2017;mittler etal.,2022)。ros主要在叶绿体、线粒体、质膜、过氧化物酶体中产生(elstner,1982;sharmaet al.,2012)。分子氧(o2)发生有氧代谢,形成具有高反应性的ros,包括含自由基的ros:羟基自由基(·oh)、超氧化物(o2·-)、有机烷氧基(ro·)和有机过氧自由基(roo·),和非自由基ros:过氧化氢(h2o2)、和单线态氧(1o2)和臭氧(o3)(dumont and rivoal,2019)。其中h2o2半衰期最长,扩散距离最远,反应具有高度选择性,是研究中最重要的ros之一(mittler,2017)。
5、活性氧的功能是依赖剂量效应的,低浓度下,ros作为体内生长和发育信号,激活基本发育过程,如根毛生长、根伸长和向性(借助生长素)、气孔闭合(借助aba)、木质素合成(借助ja)、叶形、毛状体发育、种子萌发等(suzuki et al.,2011;morales and munné-bosch,2016),同时也对植物免疫相关的信号转导过程进行微调,启动sar以促进植物防御反应发生。植物nadph氧化酶(nox),也称呼吸爆裂氧化酶同源物(rbohs),是ros代谢中重要的调控因子。据报道,gbrboh5/18和ghrbohd可激活ros产生,增强棉花对黄萎病的抗性(chang et al.,2020;huang et al.,2021)。ros过度积累则会导致氧化应激,本质上来讲氧化应激是指氧化剂和抗氧化剂之间的氧化还原反应被打破,植物体内氧化还原信号紊乱、细胞结构遭到破坏的现象(sies,2018)。有研究发现过量的ros通过改变多种蛋白质的结构和功能导致dna、rna和膜脂质发生氧化损伤,进而损害细胞功能(phua et al.,2021;mittler etal.,2022)。过表达micrornamir398b后调控ros稳态的基因被切割,棉花体内ros过度积累,对黄萎病菌的抗性降低(miao et al.,2022)。
6、为了应对氧化应激,植物已经进化出一套较为完整的由两种抗氧化剂合作清除体内多余ros维持植物氧化还原稳态的保护机制,即酶促抗氧化剂和非酶抗氧化剂(foyerand noctor,2005)。非酶抗氧化剂由低分子量化合物组成,抗坏血酸(asa)和谷胱甘肽(gsh)是其中最重要的两种,两者相较而言asa更为重要,asa的浓度通常比gsh的浓度高一千倍。非酶促抗氧化剂可以直接与ros发生反应,但在酶促抗氧化剂介导下其清除ros的效率更高。酶促抗氧化剂主要有:超氧化物歧化酶(sod)、过氧化物酶(pod)、过氧化氢酶(cat)、抗坏血酸过氧化物酶(apx)、单脱氢抗坏血酸还原酶(mdhar)、脱氢抗坏血酸还原酶(dhar)、谷胱甘肽s-转移酶(gst)、谷胱甘肽过氧化物酶(gpx)、和硫氧还蛋白(trx)等(shaban et al.,2018;phua et al.,2021)。抗氧化剂通过调控ros的含量参与植物免疫反应,gst、pod和sod被证明参与棉花感染黄萎病菌期间ros稳态的调控,因此棉花在启动防御时可以避免由ros过度积累造成的受损(li et al.,2019a;li et al.,2019b;pei et al.,2019)。目前研究发现ros水平过高或过低都不利于植物生长和抗病,找到ros最佳浓度、明确ros调控机制、把握精确调控ros水平的方法对ros在植物中更好地发挥作用至关重要。发掘调控ros的关键基因,利用生物技术精确调控该基因在棉花中表达剂量,使ros含量保持在既有利于植株生长发育又有助于抗病的范围,是抗病育种的思路之一。
7、抗坏血酸(asa),即维生素c,是一种多功能的还原剂,它包含还原型抗坏血酸(asa)、半还原型抗坏血酸(mdha)、氧化型抗坏血酸(dha)三种形式,还原型抗坏血酸和氧化型抗坏血酸合称为总抗坏血酸(total-asa)。我们目前报道的植物合成抗坏血酸的途径主要有四条:l-半乳糖途径、l-古洛糖途径、肌醇途径和d-半乳糖醛酸途径(ishikawa etal.,2018;yoshimura and ishikawa,2017)它们都以一种醛糖内酯作为抗坏血酸直接合成前体。除上述从头合成途径外,还原型抗坏血酸还通过再循环系统产生。asa是apx特定的电子供体,两个asa分子在apx催化下,将h2o2转化为h2o的同时被氧化成不稳定的单脱氢抗坏血酸(mdha)。mdha既可以自发地迅速转化为asa和dha,也可以在nad(p)h依赖性单脱氢抗坏血酸还原酶(mdar)催化下被还原成asa(foyer and shigeoka,2011)。mdar主要存在于胞质、叶绿体、过氧化物酶体、线粒体和质膜中(noctor and foyer,1998)。dha不具有抗氧化能力,以gsh为电子供体在氧化型抗坏血酸还原酶(dhar)的作用下得到电子,转化为asa(nimse and pal,2015)。gsh依赖型dhar在叶绿体、线粒体和过氧化物酶体中表达(noctorand foyer,2016)。如果不再循环,dha则发生不可逆水解,生成2,3-二酮古龙酸(moller etal.,2007)。
8、抗坏血酸在植物中主要作为抗氧化剂和酶的辅助因子发挥作用。抗坏血酸是植物最丰富的水溶性抗氧化剂(sirikhachornkit and niyogi,2010),植物过氧化物酶体、胞质溶胶、细胞核、叶绿体和线粒体中均存在较高浓度的抗坏血酸。在氧化应激中,抗坏血酸既可以直接清除体内多余的ros,又可通过asa-gsh循环帮助细胞解毒。asa-gsh循环是植物细胞主要抗氧化系统,gsh作为asa再生循环过程中dha的电子供体,在dhar催化下转化为氧化型谷胱甘肽(gssh),gssh在谷胱甘肽还原酶(gr)的催化下利用nad(p)h提供的电子可再生为gsh,不断为asa再生循环助力(foyer and noctor,2011;smirnoff and wheeler,2000)。抗坏血酸在植物应对由非生物胁迫产生的氧化应激中的作用被广泛研究。烟草和加杨的叶片因含有较多的asa,对氧化应激更耐受(foyor et al.,1995;aono et al.,1993)。镉胁迫下,多棱大麦体内的dha含量显著升高(demirevska-kepova et al.,2006)。强光照和干旱使粗枝云杉幼苗中的asa显著增加(yang et al.,2008)。紫外胁迫显著增加了金合欢幼苗的asa和dha的含量和gsh/gssg比率(agarwal et al.,2007)。有趣的是,一些研究与上述结果相反,镉胁迫下大豆根和根瘤中的asa降低(balestrasse et al.,2001);镉胁迫还分别降低了拟南芥、黄瓜和豌豆叶片中的asa含量(skorzynska-polit et al.,2003;zhang etal.,2003;romero-puertas et al.,2007),而加杨根中asa含量不受胁迫影响(romero-puertas et al.,2007)。除了对非生物胁迫下植物体内的抗坏血酸含量变化关注外,许多研究围绕asa-gsh循环过程中的apx、mdar、dhar、gr的变化展开,大部分结果认为酶反应的活跃有助于植物抵御逆境(chen etal.,2005;ushimaru et al.,2006)。总结来讲,目前多数研究认为抗坏血酸含量增加有利于植物应对氧化应激,但抗坏血酸在植物氧化应激中更全面的研究还待继续展开。另外抗坏血酸在植物应对生物胁迫时也可能发挥一定的作用,目前相关报道较少。除了作为抗氧化剂,抗坏血酸还作为辅助因子参与多个反应,例如:作为多种生物合成途径和叶黄素循环中的辅助因子,保护植物免受过量激发能量的负面影响、维持能量耗散、调节气孔功能等(niyogi,1999;smirnoff,2000)。
9、细胞色素b561(cytochrome b561,cyb561/cyb561a1)是一类包含cyb561核心结构域的跨膜家族蛋白(asard et al.,2001)。cyb561结构域由四个跨膜α-螺旋束组成,结构域内含有4个保守his残基和两个分别位于膜两侧的血红素基团,它们在细胞色素b561行使功能时发挥重要作用,家族基本结构特征保守表明这些蛋白的作用模式和生理功能可能非常相似(asard et al.,2013)。cyb561家族根据是否含有多巴胺β-单加氧酶n-末端(domon)额外结构域,分为简单cyb561s成员和cybdoms成员(tsubaki et al.,2005;asard et al.,2013)。cyb561s最初在哺乳动物分泌儿茶酚胺的囊泡中发现,后被发现在植物中广泛存在(flatmark and terland,1971;asard et al.,1989;askerlund et al.,1989;asard etal.,2000)。在植物中目前鉴定到拟南芥基因组中存在16个cyb561家族成员,二倍体雷蒙德氏棉中有20个cyb561家族成员,四倍体陆地棉中有cyb561家族成员34个,水稻中有11个cybdom家族成员(tsubaki et al.,2005;曾建国,2019;deng et al.,2023)。cyb561家族基因在植物中的数量远多于在动物,暗示着cyb561家族基因可能在植物体内有着重要且广泛的功能。
10、在动物中,cyb561s主要作为跨膜电子传递体帮助asa跨膜传递电子,促进膜另一侧mdha再生为asa以及为fe3+跨膜还原传递电子等(vargas et al.,2003;bérczi et al.,2005;bérczi et al.,2013)。牛嗜铬颗粒膜中的cyb561参与跨膜asa再生,维持asa依赖的儿茶酚胺生物合成(njus et al.,1987)。小鼠十二指肠上皮细胞细胞膜中cyb561,响应缺氧和缺铁(su and asard,2006)。植物中的cyb561s有着与动物cyb561s相似的功能(asardet al.,1989;askerlund et al.,1989)。拟南芥液泡膜上的tcytb/cyb561b1具有与动物cyb561s相似的结构特征和生理生化特征,经预测其内部含有asa结合位点,同时具有降低液泡腔中fe3+浓度的功能(asard et al.,2013);拟南芥液泡膜上的cyb561a可作为asa依赖性可逆跨膜mdha氧化还原酶发挥作用(gradogna et al.,2023);菜豆cyb561s可能是位于质膜(pm)的跨膜电子传递体,协助胞外asa再生,帮助植物应对气态氧化剂(臭氧、nox)造成的氧化应激(asard et al.,1992);关于大豆中cyb561s和玉米cyb561b1的研究认为,结构域内血红素附近带正电荷的残基可能促进asa和mdha的相互作用(bursey and poulos,2000;lad et al.,2002;nakanishi et al.,2009)。大豆pm上的cybdom介导电子从细胞内电子供体asa到细胞外电子受体fecn或铁螯合物fenta(picco et al.,2015)。植物蛋白质组学认为质膜cybdoms蛋白在某些情况下与脂筏相关(sun et al.,2008;lefebvre etal.,2007)。综上,大多数研究认为植物细胞色素b561家族基因主要功能与动物细胞色素b561家族基因相同,均为参与asa跨膜电子传递、参与mdha再生asa过程和参与fe3+跨膜转运,但该家族基因在棉花中的功能未见报道。
11、植物细胞色素b561家族基因参与植物应对非生物胁迫的报道较少,西瓜质膜上cyb561b在干旱和高光胁迫下上调表达(nanasato et al.,2005);水稻中位于pm和内质网的oscybdomg1在水稻耐盐、植株生长和产量的调节中发挥着重要作用,突变该基因使植物体内asa含量和脱氢抗坏血酸比率(asa/dha)降低,进而导致h2o2积累、植物耐盐性降低、植物生长和粮食产量严重受到影响(deng et al.,2023),细胞色素b561家族基因参与植物应对生物胁迫未见报道。
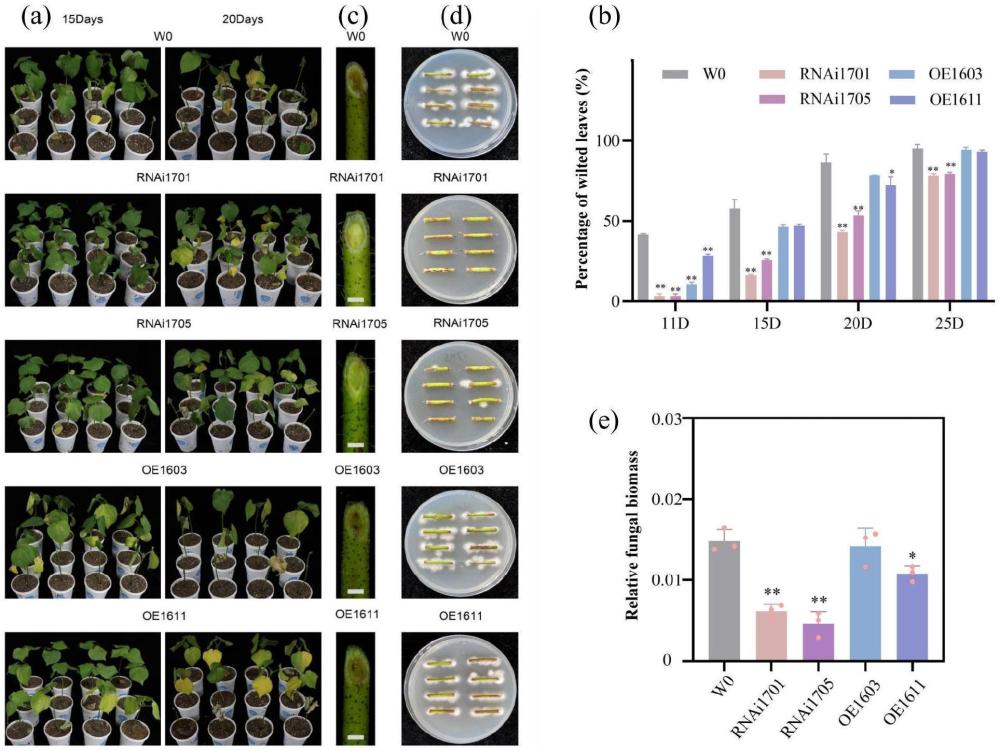
12、本实验室通过黄萎病菌诱导后棉花根部转录组测序结果,发现参与抗坏血酸再生过程的基因ghvdr1,受黄萎病菌诱导后表达水平显著下调;而在棉花中瞬时沉默该基因后,植株的抗病性显著增强。进一步以异源四倍体陆地棉材料w0为受体创制了稳定的ghvdr1下调表达材料和过表达材料。该基因干扰表达和过表达对棉花的纤维产量和品质没有显著影响。ghvdr1基因编码细胞色素b561蛋白(cyb561),位于棉花第11号部分同源染色体上,a/d亚组各一个拷贝,分别为gh_a11g1488和gh_d11g1514,同源性高。通过接菌黄萎病抗性鉴定,结果表明,ghvdr1干扰植株具有显著提高的黄萎病抗性,过表达对棉花抗病性没有显著影响。
13、棉花黄萎病危害巨大,治理困难,而占全球种植面积90%以上的陆地棉抗源匮乏。鉴于此,ghvdr1的发现及抗病分子机理研究,可以拓宽对棉花黄萎病抗性的认识。通过调控目标基因的表达,实现精确调控棉花生长和抗病之间的平衡,培育抗性显著提高的新种质并在生产上应用,为棉花抗黄萎病育种提供理论和材料基础。
技术实现思路
1、本发明的目的是提供棉花ghvdr1基因或与ghvdr1基因相关的生物材料在提高棉花黄萎病抗病性或培育黄萎病抗病性提高的棉花新种质中的应用。以此基因为靶基因,通过rnai干扰技术证明了干扰表达ghvdr1可以显著提高植株对黄萎病的抗性。以此基因为靶基因,培育生长发育正常,棉纤维产量、品质与对照无显著差异的沉默、干扰或抑制所述棉花ghvdr1基因表达的生物材料,实现其在提高棉花抗病性或培育黄萎病抗病性提高的棉花新种质中的应用。
2、本发明的另一目的在于提供一种提高棉花抗黄萎病的方法。
3、本发明的又一目的在于提供一种棉花ghvdr1基因,并提供该基因在陆地棉品种tm-1的中的全长cdna orf核苷酸序列及编码的氨基酸序列。
4、本发明的目的通过以下技术方案实现:
5、第一方面,本发明请求保护如seq id no.1所示的棉花cyb561家族ghvdr1基因或与ghvdr1基因相关的生物材料在如下(a1)或(a2)中的应用:
6、(a1)提高棉花黄萎病抗病性;
7、(a2)培育黄萎病抗病性提高的棉花新种质。
8、进一步,所述的与ghvdr1基因相关的生物材料为如下(b1)或(b2)或(b3)所述的生物材料:
9、(b1)所述ghvdr1基因编码的蛋白ghvdr1;
10、(b2)含有所述ghvdr1基因的生物材料;
11、(b3)用于沉默、干扰或抑制所述ghvdr1基因的生物材料。
12、更进一步,(b1)中所述ghvdr1基因编码的蛋白具有如seq id no.2所示的氨基酸序列。
13、更进一步,(b2)中所述的含有所述ghvdr1基因的生物材料为如下(c1)~(c6)中的至少一种:
14、(c1)含有所述ghvdr1基因的表达盒;
15、(c2)含有所述ghvdr1基因的重组载体、或含有(c1)所述表达盒的重组载体;
16、(c3)含有所述ghvdr1基因的重组微生物、或含有(c1)所述表达盒的重组微生物、或含有(c2)所述重组载体的重组微生物;
17、(c4)含有所述ghvdr1基因的转基因植物细胞系、或含有(c1)所述表达盒的转基因植物细胞系、或含有(c2)所述重组载体的转基因植物细胞系;
18、(c5)含有所述ghvdr1基因的转基因植物组织、或含有(c1)所述表达盒的转基因植物组织、或含有(c2)所述重组载体的转基因植物组织;
19、(c6)含有所述ghvdr1基因的转基因植物器官、或含有(c1)所述表达盒的转基因植物器官、或含有(c2)所述重组载体的转基因植物器官。
20、更进一步,(b3)中所述用于沉默、干扰或抑制所述ghvdr1基因的生物材料为如下(d1)~(d4)中的至少一种:
21、(d1)所述ghvdr1基因的干扰片段或沉默片段;
22、(d2)用于扩增(d1)所述ghvdr1基因干扰片段或沉默片段的引物;
23、(d3)所述ghvdr1基因的干扰表达载体或沉默载体;
24、(d4)含有如(d3)所述干扰表达载体或沉默载体的重组微生物。
25、进一步,上述的应用为通过抑制棉花ghvdr1基因的表达量或降低棉花ghvdr1基因编码的蛋白ghvdr1的水平(活性或含量)提高棉花抗病性。更进一步,所述的抑制棉花ghvdr1基因的表达量的过程为:构建该基因的干扰表达载体,通过农杆菌介导法将构建的干扰表达载体转化棉花,获得ghvdr1基因表达显著降低的棉花材料。
26、第二方面,本发明请求保护一种提高棉花抗病性的方法,通过抑制所述棉花ghvdr1基因的表达量或降低由棉花ghvdr1基因编码的蛋白ghvdr1的水平(活性或含量)提高棉花黄萎病抗病性。
27、第三方面,本发明请求保护一种棉花细胞色素b561家族的ghvdr1基因,该基因具有如seq id no.1所示核苷酸序列。
28、第四方面,本发明请求保护由所述的ghvdr1基因编码的蛋白,该蛋白具有如seqid no.2所示的氨基酸序列。
29、第五方面,本发明请求保护与所述棉花ghvdr1基因相关的生物材料,该生物材料为含有所述ghvdr1基因的表达盒、重组载体、转基因细胞系或重组菌;或用于沉默、干扰或抑制所述棉花ghvdr1基因的生物材料;具体为含有所述棉花ghvdr1基因的干扰或沉默特异性片段的重组载体、表达盒和重组菌。
30、本发明克隆一种棉花细胞色素b561家族的ghvdr1基因,该基因的orf序列如seqid no.1所示。该基因编码的蛋白具有如seq id no.2所示的氨基酸序列。研究发现,抑制所述ghvdr1基因的表达能显著提高棉花对黄萎病的抗性。将目标基因的干扰或沉默表达载体导入现有的推广品种中,对推广品种进行棉花抗病性改良。本发明的具体实施方式中,以所述的棉花ghvdr1基因为靶基因,通过基因干扰方法,抑制ghvdr1基因表达,培育棉花抗病性显著改良的新种质并在生产上应用。
31、本发明的优点表现在:
32、(1)首次对棉花中的ghvdr1基因进行系统的序列结构、表达模式及功能分析,明确ghvdr1在棉花生长发育及抗病性中的重要作用。
33、细胞色素b561在植物基因功能研究中被广泛报道参与抗坏血酸再生和fe3+还原,有报道称其可以通过调节抗坏血酸的含量影响植株体内ros,进而参与植物抵御非生物胁迫,其在植物抗病中未见报道。本发明所克隆的ghvdr1基因,前人未曾研究过其在棉花中功能。我们通过系统的分子生物学实验证实了棉花中ghvdr1表达水平的下降对植株产量和纤维品质影响较小。干扰表达ghvdr1植株通过降低抗坏血酸再生关键基因表达量,降低抗坏血酸含量和抗坏血酸氧化还原效率,增加h2o2的含量,增强植株抗病性。研究结果揭示了棉花中ghvdr1调控植物生长发育及黄萎病抗性的分子机制,也为进一步探究植物如何精准平衡生长-免疫的研究方向提供了新思路与参考。
34、(2)首次对喷施抗坏血酸的植株进行抗病性鉴定,明确抗坏血酸在棉花抗病性中的重要作用。
35、外施抗坏血酸后野生型陆地棉w0和与野生型相比具有较强抗病性的干扰表达ghvdr1植株的抗病性,与喷施水的植株相比显著增强,表明抗坏血酸外施降低植株抗病性。这一结果既为ghvdr1抗病机理解析提供进一步补充,又为棉花抗病分子育种提供新思路。
本文地址:https://www.jishuxx.com/zhuanli/20241009/305956.html
版权声明:本文内容由互联网用户自发贡献,该文观点仅代表作者本人。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如发现本站有涉嫌抄袭侵权/违法违规的内容, 请发送邮件至 YYfuon@163.com 举报,一经查实,本站将立刻删除。